
Abstract
Apoptosis is a form of regulated cell death (RCD) that involves proteases of the caspase family. Pharmacological and genetic strategies that experimentally inhibit or delay apoptosis in mammalian systems have elucidated the key contribution of this process not only to (post-)embryonic development and adult tissue homeostasis, but also to the etiology of multiple human disorders. Consistent with this notion, while defects in the molecular machinery for apoptotic cell death impair organismal development and promote oncogenesis, the unwarranted activation of apoptosis promotes cell loss and tissue damage in the context of various neurological, cardiovascular, renal, hepatic, infectious, neoplastic and inflammatory conditions. Here, the Nomenclature Committee on Cell Death (NCCD) gathered to critically summarize an abundant pre-clinical literature mechanistically linking the core apoptotic apparatus to organismal homeostasis in the context of disease.
Similar content being viewed by others


Facts
-
Intrinsic and extrinsic apoptosis are forms of regulated cell death (RCD) promoting the cellular demise along with the activation of proteases of the caspase family.
-
In mammalian organisms, executioner caspases are activated after cells are already committed to die.
-
Apoptosis can be manipulated by genetic or pharmacological means, and multiple genetically engineered animal models and pharmacological tools to modulate apoptosis have been developed.
-
Apoptosis is intimately involved in both (post-)embryonic development and adult tissue homeostasis.
-
Apoptosis deregulation promotes oncogenesis and contributes to the etiology of multiple human disorders, including cardiovascular, hepatic, renal, inflammatory and neurological conditions.
-
To date, venetoclax is the only apoptosis inducer that has received regulatory approval for use in humans.
Open Questions
-
Will inhibitors of apoptotic caspases with elevated target specificity become available?
-
Will agents specifically conceived to modulate apoptosis enter the clinical practice to treat solid tumors or other human disorders beyond hematological malignancies?
-
Is it conceivable to design combinatorial strategies aimed at inhibiting apoptosis while interrupting compensatory activation of other RCD signaling cascades?
-
Will it be possible to specifically inhibit apoptotic signaling without impacting on other processes dependent on apoptosis regulators such as differentiation, proliferation, and inflammatory reactions?
Introduction
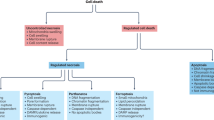
The health and homeostasis of multicellular organisms depend on the tight balance between cell proliferation and cell death. In this context, a large body of experimental evidence has demonstrated the existence of a form of regulated cell death (RCD) that is executed by a genetically programmed process, and hence amenable to manipulation by genetic or pharmacological means [1]. Over the past decades, multiple variants of RCD have been characterized at the genetic, biochemical, functional, and immunological level [2,3,4,5,6,7,8]. For instance, programmed cell death (PCD) has been functionally defined as a modality of RCD activated under purely physiological conditions (i.e., in the absence of perturbations of extracellular or intracellular homeostasis) in the context of embryonic/post-embryonic development or adult tissue homeostasis [1, 9]. Conversely, pathological RCD is invariably initiated in the context of failure to adapt to shifts in extra-cellular or intra-cellular homeostasis, constituting a de facto organismal program for the elimination of excessively damaged and/or potentially harmful cells, such as cells infected with pathogens [1, 10]. From a biochemical perspective, an increasing number of RCD modalities have been defined by the Nomenclature Committee on Cell Death (NCCD) based on the mechanistic involvement of specific molecular components [1, 11]. For instance, apoptotic cell death has been defined as a form of RCD that is promoted by proteases of the caspase family, namely caspase 3 (CASP3), CASP6 and CASP7, and initiated by CASP8 and CASP9 [1, 12, 13]. However, in mammalian organisms, with the exception of CASP8, apoptotic caspases simply accelerate RCD because their activation occurs when cells are already committed to die [1, 14,15,16]. This means that contrarily to simpler organisms (e.g., Caenorhabditis elegans), in which apoptotic caspase elimination fully rescues cells from death, in mammals, apoptotic cell death can at most be retarded but not prevented by pharmacological or genetic strategies inhibiting the activity of these caspases. Mitochondrial permeability transition (MPT)-driven necrosis, necroptosis, ferroptosis, pyroptosis, parthanatos, entotic cell death, NETotic cell death, lysosome-dependent cell death, and autophagy-dependent cell death represent forms of RCD that involve precise molecular events and hence can also be manipulated with pharmacological or genetic interventions [1,2,3,4,5,6, 17,18,19]. Other RCD modalities have been recently identified, such as alkaliptosis [20], cuproptosis [21] and PANoptosis (involving the simultaneous activation of pyroptosis, apoptosis, and necroptosis) [22], and their signal transduction modules are under investigation. The importance of these latter forms of RCD in health and disease is not yet known.
Along with the identification of key RCD regulators and the advent of modern tools for genetic manipulation, a great experimental effort has been devoted to elucidating the role of RCD in the physiopathology of multi-cellular organisms [23]. Thus, various studies in animals (mostly rodents) genetically altered to lack or over-express components of the apoptotic apparatus (either at the whole-body level or in selected cell/tissue types) have provided formal proof of the relevance, but not always the exquisite requirement, of apoptosis for embryonic and fetal development or adult tissue homeostasis [24,25,26].
Along similar lines, pharmacological and genetic tools aimed at altering apoptotic signaling in pre-clinical disease models revealed the mechanistic contribution of apoptosis to the etiology of various conditions associated with the loss of post-mitotic or (in certain settings) non-post-mitotic cells, including a panel of neurological, cardiovascular, renal, hepatic, and inflammatory disorders [24]. Extensive studies over the last five decades highlighted the apoptotic machinery as a major target for the development of new therapeutic interventions [27], not only for the induction of cell death in the context of disrupted tissue homeostasis (e.g., for neoplastic diseases) [28], but also for the inhibition of cell death in the context of ischemic, degenerative and inflammatory conditions [29, 30]. However, while at least one drug designed to induce apoptosis is currently approved for use in humans, namely the BCL2 apoptosis regulator (BCL2) inhibitor venetoclax [31,32,33,34], which is used alone or in combinatorial regimens for the treatment of chronic lymphocytic leukemia (CLL), small lymphocytic lymphoma and acute myeloid leukemia (AML) [31, 35,36,37,38], no other agents specifically conceived to inhibit the apoptotic apparatus have been licensed for clinical practice so far. The broad-spectrum caspase inhibitor emricasan received fast-track designation by the US Food and Drug Administration (FDA) for the treatment of non-alcoholic steatohepatitis in 2016 but demonstrated inconsistent clinical efficacy [39,40,41], and – as of now – is not approved for therapy in humans.
The lack of clinically approved, selective apoptosis inhibitors and the inconclusive performance of emricasan in recent trials reflect several aspects of (apoptotic and non-apoptotic) RCD that began to emerge only recently (Fig. 1). First, while detecting cell death as well as biomarkers of specific RCD variants in vitro is relatively straightforward [42], precise quantification of cell death in vivo in adult tissue remains challenging, at least in part because of rapid disposal of cell corpses by efferocytosis [43,44,45,46]. Thus, the actual contribution of cell death to the etiology of various human disorders is difficult to quantify by observational approaches [47, 48]. Second, while for a long-time, specific forms of RCD were considered virtually independent entities, it recently became clear that the molecular machinery for RCD is composed of highly interconnected modules characterized by substantial redundancy, backup pathways and feedback loops [10, 49, 50]. Thus, molecules that inhibit one specific form of RCD may ultimately be unable to confer actual cyto- and tissue protection instead only altering the kinetic and biochemical manifestations of death by allowing the engagement of a different RCD sub-routine. For instance, while CASP8 is a major signal transducer in death receptor (DR)-driven apoptosis (see below), it intrinsically inhibits necroptosis induced by DRs and other signaling pathways, such as Toll-like receptor (TLR) signaling [51,52,53], suggesting that caspase inhibition in the context of DR signaling may promote necroptotic cell death [54,55,56,57]. Together with a low target specificity and selectivity within the caspase family [57], this can explain the inadequate efficacy of emricasan observed in pre-clinical and clinical studies. Third, even in the hypothetical scenario of agents capable of simultaneous inhibition of all (known and unknown) RCD pathways, loss of cellular homeostasis due to failing adaptation to stress generally involve degenerative processes that at some stage cannot be reversed, such as widespread mitochondrial permeabilization and loss of RNA and protein synthesis [4, 58,59,60], i.e., even if all RCD modalities could be blocked effectively, cells might undergo uncontrolled necrotic death. In this setting, cell death may occur as a consequence of an irremediable degeneration of cellular functions that can no longer be rescued pharmacologically or even genetically [61]. Supporting these latter notions, accumulating literature indicates that, at least in mammalian systems, perhaps with the exception of CASP8, so-called apoptotic caspases mainly control the kinetics of apoptotic cell death and its immunological manifestations, but not whether cell death ultimately occurs or not [15, 16]. This points to the caspase family as a major regulator of organismal homeostasis via control of inflammatory responses [62, 63]. The simultaneous inhibition of multiple caspases, as for instance by emricasan, may thus also impact inflammation, as was demonstrated for tumor necrosis factor (TNF)-induced systemic inflammatory respiratory syndrome (SIRS) in vivo for the pan caspase-inhibitor zVAD-fmk [54, 64]. To complicate matters, multiple components of the core apoptotic machinery, including caspases and multiple members of the BCL2 family have been reported to regulate a variety of non-apoptotic functions beyond inflammation, such as mitochondrial energy production, Ca2+ signaling and terminal differentiation [65,66,67,68,69,70,71,72]. Structurally, distinguishing between apoptotic and non-apoptotic functions of caspases and the BCL2 family remains challenging. Finally, there is a hitherto unclarified heterogeneity in the regulation of RCD at distinct anatomical sites (possibly linked to micro-environmental features) at distinct stages of cellular differentiation, and in the context of diverse patho-physiological states (e.g., in young vs. adult and aged individuals).

The clinical development and success of agents inhibiting apoptosis is limited by multiple contributory causes, including potential non-apoptotic, accessory or even protective roles of the targeted proteins (exemplified by the involvement of certain BCL2 family members, caspases and death receptors in processes as diverse as inflammation, cell differentiation, cell proliferation and cell survival), the high interconnectivity between RCD pathway (potentially leading to the activation of compensatory RCD variants in response to the inhibition of a specific RCD type), the low specificity and selectivity of the inhibitors developed so far (exemplified by the broad-spectrum caspase inhibitors) and the difficulty to precisely determine and quantify cell death in vivo. RCD regulated cell death.
All these issues should also be kept under consideration in the context of the present review, in which the NCCD aims at critically discussing a large amount of pre-clinical data in support of a key role for the apoptotic machinery in mammalian diseases. Specifically, the interpretation of results of genetic and pharmacological experiments presented herein should place particular attention on the aforementioned connectivity amongst different RCD variants as well as on discriminating between essential vs. accessory aspects of cell death [14]. Another issue to be considered is the fact that most conclusions are based on use of knockout/congenic mice which often present other passenger mutations potentially influencing the observed phenotype [73]. Our objective is not only to provide a critical summary of the existing literature, but also to offer an updated framework for interpretation of these findings in view of currently accepted models of RCD signaling.
Intrinsic apoptosis in disease
There are substantive supporting data from genetic studies to demonstrate that the molecular machinery for intrinsic apoptosis (described in Box 1 and Fig. 2) is involved in embryonic and fetal development as well as in adult tissue homeostasis. Numerous preclinical studies in animal models of disease demonstrate that intrinsic apoptosis contributes to etiology in various disorders involving the loss of not only post-mitotic, but also non-post-mitotic tissues, including neurological, cardiac, renal, hepatic, autoimmune/inflammatory, oncological, and infectious conditions. However, as discussed above, the interpretation of these results should be taken with caution given the high interconnectivity of RCD pathways and the crosstalk between RCD and inflammatory response. Moreover, the activation of executioner caspases occurs after cells are already committed to intrinsic apoptosis [15, 16]. Accordingly, caspase inhibition only delays the execution of cell death. In this context, the phenotypes observed under apoptotic caspase-deleted or inhibited conditions may reflect cell-extrinsic effects of caspase activity such as the release of immunomodulatory and cytotoxic signals from dying/dead cells, including damage-associated molecular patterns (DAMPs) or cytokines (this concept is extensively discussed in [14]). These phenotypes may also stem from the lack of processes independent of intrinsic (or extrinsic) apoptosis, as, for instance, the lack of CASP3-mediated cleavage of gasdermin E (GSDME) leading to impaired pyroptosis and associated inflammatory response [74, 75].

Intrinsic apoptosis can be activated by a range of extracellular or intracellular stimuli, including, but not limited to, DNA damage, endoplasmic reticulum (ER) or oxidative stress, growth factor withdrawal or microtubular alterations. The critical step of the intrinsic apoptosis is the activation of the pro-apoptotic effectors of the BCL2 family, BAX, BAK and possibly BOK, which drives the outer membrane permeabilization (MOMP) and commits cells to death. MOMP results in the release from the mitochondrial intermembrane space into the cytosol of proapoptotic proteins, including CYCS and SMAC. CYCS assembles with APAF1, dATP and pro-CASP9 into the apoptosome, leading to the activation of CASP9, which in turn promotes the activation of the executioner caspases CASP3 and CASP7. The activation of the executioner caspases is facilitated by SMAC, which sequesters and/or degrades members of IAP family that inhibit apoptosis.
Below, we will provide details of the pro-apoptotic BCL2 proteins, the anti-apoptotic BCL2 proteins, the components of the apoptosome—a platform for the activation of initiator caspases composed of cytochrome c, somatic (CYCS), apoptotic peptidase activating factor 1 (APAF1) and pro-CASP9—and effector caspases in disease. The instances of involvement encompass participation in the pathogenic mechanisms as well as experimental deletion or inhibition as a means of exploring potential utility as treatment targets. The effects of these regulators and effectors of the intrinsic apoptosis pathway on health are described in Box 2, Box 3 and Box 4.
Neurological disorders
Intrinsic apoptotic factors are implicated in the pathophysiology of numerous neurological diseases (Fig. 3). In a mouse model of amyotrophic lateral sclerosis (ALS), deletion of BCL2-associated X protein (Bax) reduces neuronal cell death coupled to attenuated motor dysfunction and neuromuscular degeneration [76]. Additional ablation of BCL2-antagonist/killer 1 (Bak1) further enhances neuroprotection, resulting in improved overall animal survival [77]. Similar protective effects were observed in mice lacking the BH3-only proteins BCL2 like 11 (BCL2L11, best known as BIM) and BCL2 binding component 3 (BBC3, best known as PUMA), as well as in transgenic mice overexpressing BCL2, X-linked inhibitor of apoptosis (XIAP) [78,79,80,81,82]. Moreover, intra-cerebroventricular administration of the broad-spectrum inhibitor Z-VAD-FMK protects mice from ALS [83], although whether such protection arises from the inhibition of intrinsic apoptosis remains to be formally established. Bax deletion also attenuates neuromuscular dysfunctions in a mouse model of congenital muscular dystrophy (another neurodegenerative disease affecting motoneurons) [84], while BCL2 overexpression limits neuromuscular disease progression in some (but not all) mouse models of progressive motor neuronopathy and muscular dystrophy [85,86,87]. Finally, genetic or pharmacological inhibition of poly (ADP-ribose) polymerase family, member 1 (PARP1) and PARP2 halts axonal degeneration and improves related motor phenotypes in C. elegans models of ALS [88].

Intrinsic apoptosis is directly or indirectly involved in the pathogenesis of multiple neurological disorders, including neurodegenerative diseases, brain damage caused by traumatic injury or neurotoxicity as well as neuromuscular and retinal disorders.
Multiple components of the molecular machinery for intrinsic apoptosis, including BAX, PUMA, BH3 interacting domain death agonist (BID), Harakiri, BCL2 interacting protein (contains only BH3 domain) (HRK), were shown to drive neuronal death in Alzheimer’s disease (AD) and Parkinson’s disease (PD) models [89,90,91,92,93,94,95,96,97,98,99,100,101]. Thus, overexpression of BCL2 decreases the appearance of early pathological markers of AD, such as amyloid precursor protein (APP) and microtubule-associated protein tau (MAPT, best known as tau) cleavage, which depend on caspases [102,103,104], resulting in attenuated neurological defects [105, 106]. Some findings indicate a role of apoptotic caspases in the pathogenesis of AD. However, as discussed above, during intrinsic apoptosis, caspases simply accelerate the course of cell death, and, so, such effects may be linked to the release of cytotoxic and pro-inflammatory factors from dying cells. In more detail, pharmacological inhibition of CASP3 reduces early synaptic failure in mouse models of AD, ultimately improving cognitive defects [107]. Moreover, expression of a mutated form of amyloid β (an APP cleavage product) or administration of broad-spectrum caspase inhibitors attenuates synaptic defects in models of AD, an effect only partially recapitulated by CASP3-specific inhibitors [108]. Along similar lines, deletion of Casp2 was reported to provide protection from synaptic loss and cognitive decline in a mouse model of AD [109]. Such protection may be linked to the generation of a specific tau cleavage product (∆tau314) by CASP2, which is reported to impair cognitive and synaptic function by promoting the missorting of tau to dendritic spines [110, 111]. Accordingly, CASP2 inhibitors blocked tau truncation and restored excitatory neurotransmission in mouse models of tauopathies, including AD [112, 113]. A role for CASP4 in AD pathogenesis has also been reported [114, 115]. Moreover, studies using the senescence-accelerated OXY5 rat model of AD demonstrated that the treatment with mitochondria-targeted antioxidant SkQ1 improved mitochondrial fitness and slowed down the signs of Alzheimer’s disease-like pathology in older rats [116]. Lack of BIM (due to deletion of Bcl2l11) also confers protection to dopaminergic neurons in experimental PD imposed by inhibition of mitochondrial complex I, an effect that depends on BAX activation [117]. In addition, genetic deletion or down-regulation of Casp3, as well CASP3 inhibition by transgenic, neuron-restricted expression of XIAP, protects mice against pharmacologically induced PD, attenuating both dopaminergic neuron alterations and behavioral deficits [118,119,120,121]. Whether protection arises from the lack of cell-intrinsic or cell-extrinsic processes dependent on apoptotic caspases has not been investigated. Finally, pharmacological inhibition of CASP3 confers neuroprotection in a rat model of Huntington’s disease (HD) [122,123,124]. That said, the precise mechanisms whereby components of the molecular apparatus for intrinsic apoptosis influence neurodegeneration need to be further explored. Two studies in clear contradiction to each other reported that, at sublethal doses, pharmacological inhibition of myeloid cell leukemia sequence 1 (MCL1) improved disease outcome in a mouse model of AD with a mechanism independent of apoptosis induction and involving the stimulation of mitophagy [125], but that Mcl1 haploinsufficiency accelerated the degeneration and dysfunctionality of motor neurons in mice [126]. Also, there is evidence that necroptosis or ferroptosis rather than apoptosis can be the major contributor in neuronal cell destruction during AD [127, 128]. Finally, although Bax deletion prevents the demise of cerebellar granule neurons in a transgenic model of inherited prion disease [129], the direct contribution of BAX to neurotoxicity during prion disorders remains a matter of controversy [130].
BCL2 family proteins have also been reported to contribute to axonal degeneration and neuronal cell death in animal models of brain trauma, degeneration, or neurotoxicity [131,132,133]. Thus, BAX- or BID-deficient mice, as well as transgenic mice overexpressing BCL2, display increased survival of cortical or hippocampal neurons after experimental traumatic brain injury, as compared to wild-type mice [134,135,136,137]. Moreover, transgenic BCL2 overexpression protects mouse neurons against the detrimental effects of transection of the sciatic nerve [138]. Likewise, BAX deficiency enhances the survival of oligodendrocytes in mice subjected to spinal cord injury [139]. Both neuroprotection and functional improvements were observed in rat or mouse models of traumatic spinal cord injury upon local administration of Z-VAD-FMK) and other caspase inhibitors [140,141,142]. However, these findings need to be validated given the low selectivity of these inhibitors among caspases. Of note, in rats, post-traumatic neuroprotection can further be improved by combined inactivation of PARP1 and CASP3 [143], suggesting a potential involvement for PARP1-dependent parthanatos in the process.
Deletion of Bax (but not the genes encoding BIM, PUMA or BID), as well as Bax haploinsufficiency, prevents the death or degeneration of retinal ganglion cells in mice subjected to optic nerve injury [144,145,146,147]. Moreover, the demise of injured retinal ganglion cells is exacerbated in mice with a conditional loss of Bcl2l1 (leading to lack of BCL-XL) [148] and decreased in transgenic mice over-expressing XIAP [149] or BCL-XL [150] in the eye, as well as in rodents treated with an XIAP-derived cell-permeant peptide targeting CASP9 [151], or a CASP3-targeting small-interfering RNA (siRNA) [152, 153]. Moreover, transgenic or adenovirus-driven XIAP expression protects the retina in various animal models of retinal disease, degeneration, or ischemia [154,155,156,157,158,159], while a BCL-XL inhibitor alleviates pathogenic neo-vascularization during diabetic retinopathy [160]. Genetic deletion of Casp9 from endothelial cells protected retinal ganglion cells from ischemic death, supporting non-cell autonomous functions of CASP9 [151]. Of note, CASP7 seems to play a crucial role in retinal ganglion cell death, as demonstrated in a model of optic injury in Casp7−/− mice [161]. However, both pro-survival (BCL2) and pro-apoptotic (BAK1, BAX and BIM) BCL2 family members contribute to retinal neo-vascularization in response to experimental ischemic retinopathy [162,163,164]. In one of these models, such an effect was linked to an increased survival of endothelial cells in the absence of BAX and BAK1 [164]. Persistent endothelial cells promote indeed rapid tissue re-vascularization, thus preventing the occurrence of a pathogenic excessive neovascularization. Moreover, the inhibition of the intrinsic apoptotic pathway by c-Jun N-terminal kinase 1 (Jnk1) deletion or the administration of a broad-spectrum caspase inhibitor led to reduced choroidal neo-vascularization in the murine model of wet age-related macular degeneration (AMD) [165]. These observations may indicate that factors released by dying cells regulate neo-vascularization in the retina or other eye tissues.
Deletion of Bax, Hrk or Casp3 as well as transgenic overexpression of XIAP prevents neuronal loss and/or axon degeneration in mouse models of trophic factor deprivation including nerve growth factor (NGF) withdrawal [166,167,168]. Conversely, lack of BIM or PUMA does not limit hippocampal neuronal injury upon experimental excitotoxicity [169, 170]. Moreover, while in vivo delivery of an XIAP fusion protein protects neurons against death induced by glutamate or kainic acid [171], kainic acid-mediated neurodegeneration cannot be rescued by the CASP3 inhibitor DEVD-CHO [172]. Conversely, BIM appears to be activated during excitotoxicity [173], and Bcl2l11−/− mice (which lack BIM) display attenuated neuro-degeneration after experimental seizures induced by administration of kainic acid into the amygdala, at least in part because of decreased neuronal cell death in the hippocampus (but not in the neocortex) [174]. Moreover, data from knockout mice suggest that experimental seizure-induced neuronal death involves BCL2-associated agonist of cell death (BAD), BCL2 interacting killer (BIK), BCL2 modifying factor (BMF), or PUMA [175,176,177,178] and that BCL2-like 2 (BCL2L2; best known as BCL-W) may provide neuroprotective, seizure-suppressive functions [179]. Confirming a certain degree of functional redundancy, phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1, best known as NOXA) and BID seem dispensable for RCD driven by excitotoxicity, as shown in kainic acid-treated animals [180, 181].
Intrinsic apoptosis is also involved in neuronal apoptosis post-ischemic injury in both developing and adult brains. In a mouse model of neonatal hypoxia-ischemia, neuroprotection was documented upon deletion of Bax [182], simultaneous absence of BIM and BAD [183], or transgenic overexpression of XIAP [184]. Conversely, Xiap−/− mice are sensitized to neonatal hypoxia-ischemia injury [185]. Apparently at odds with these findings, Casp3−/− mice display increased vulnerability to such experimental perturbation, possibly due to complementary over-activation of CASP3-independent pathways [186]. Of note, the absence of CASP3, BAX, or PUMA (but not the absence of NOXA, BIM or HRK) also confers neuro-protection to newborn mice acutely exposed to ethanol [187,188,189], while loss of BAX is neuroprotective in newborn mice exposed to isoflurane [190] as well as ionizing radiation [133, 191]. At the same time, it is interesting to note that BAX-dependent neuronal RCD also contributes to reactive microgliosis during the recovery of the developing brain from acute alcohol exposure [192], pointing to an etiological role for activation of microglial cells by dead neurons.
Bax−/− mice displayed pronounced neuroprotection when subjected to distinct experimental brain injuries, including middle cerebral artery occlusion [193]. A similar protection against experimental ischemic insults has been observed in mice deficient for BMF [194], or BID [195,196,197]. Conversely, NOXA seems to be dispensable for neuronal damage induced by experimental ischemic stroke [194]. Moreover, the absence of BID fails to protect mice from ischemia-reperfusion, although it limits the associated inflammatory response [198]. Transgenic over-expression of BCL2, BCL-XL or XIAP as well as inhibition of apoptotic caspases or genetic deletion of CASP6 ameliorates neuronal survival upon global ischemia, focal ischemia or stroke [199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215]. It should be noted, however, that in these settings neuroprotection by inhibition or deletion of caspases may be related to the lack of cell-extrinsic or apoptotic-unrelated roles of caspases. Morevoer, various examples of caspase-independent neuronal death after cerebral ischemia have been reported [216,217,218,219]. In this context, it is important to note that apoptosis is dynamically regulated during lifespan in the brain [24]. Indeed, while immature brain cells express high levels of many BCL2 proteins [133, 220, 221], most of these proteins are downregulated in the adult brain, when most post-mitotic neural cells become resistant to apoptosis [131, 222]. This may help explain the divergent findings on the mechanisms of neural cell death reported above.
Cardiovascular conditions
While a role for RCD in non-reperfused myocardial infarction remains questionable, apoptosis and other cell death programs including necroptosis, MPT-driven necrosis, ferroptosis, pyroptosis and autosis appear to contribute to cardiomyocyte death and tissue damage during myocardial infarction with reperfusion (also referred to as myocardial ischemia-reperfusion injury). However, the relative importance of the specific RCD mode and how it interconnects mechanistically and functionally with other RCD pathways to produce an integrated response remains poorly understood. For example, Bak1−/−mice with a cardiomyocyte-specific deletion of Bax displayed considerably reduced infarct size as compared to their wild-type littermates when subjected to experimental myocardial ischemia-reperfusion, although it remains unclear whether these effects are attributable to reductions in apoptosis or MPT-driven necrosis [223,224,225], a RCD variant shown to participate in the pathogenesis of ischemic stroke [226]. Protection against myocardial ischemia-reperfusion has also been reported in transgenic mice overexpressing BCL2 [227,228,229] or a BCL-XL-derived peptide [230]. Likewise, deletion of Bbc3 (leading to lack of PUMA) ameliorates myocardial ischemia-reperfusion injury [231], ultimately translating into increased survival [232]. Moreover, neurotrophin-3 was reported to confer cardioprotection fromischemic and reperfusion injuries by reducing BIM levels [233]. Broad spectrum caspase inhibition [234,235,236] and XIAP mimicking peptides [237] were shown to modestly reduce myocardial infarct size. Finally, simultaneous deletion of Casp3 and Casp7 had no cardioprotective effect during reperfused myocardial infarction [238], in line with the notion that the absence of caspases only delays cell death.
In contrast to the large burst of cell death over several hours characterizing myocardial infarction, cardiomyocytes are lost gradually over months to years during heart failure with reduced ejection fraction [3]. The role of intrinsic apoptosis in these heart conditions is, however, debated. In a mouse model of cardiomyopathy based on the deletion of desmin (Des), the cardiomyocyte-specific over-expression of BCL2 reduces cardiac lesions and hypertrophy coupled to ameliorated cardiac functionality [239]. However, despite improved survival, these mice show increased levels of necrosis due to the activation of alternative cell death pathways [240]. Moreover, Casp3−/− mice display enhanced vulnerability to experimental cardiomyopathy, at least in part reflecting the inefficient activation of pro-survival AKT serine/threonine kinase 1 (AKT1) signaling [241]. As an alternative explanation, the absence of CASP3 may foster RCD-driven inflammation associated with increased type I interferon (IFN) release [242,243,244]. Indeed, experimental data linking dysregulated type I IFN release and cardiac conditions have recently emerged [245].
As for therapeutic interventions, cardioprotective effects have been achieved by inhibition of CASP3 in rodent models of myocardial dysfunction induced by endotoxin [246], burn injury [247] or hypoxia [248], although perhaps such effects can be attributed to the lack of cell-extrinsic or apoptosis-unrelated effects of caspase activity. Moreover, inhibition of BAX prevents cardiotoxicity induced by doxorubicin in zebrafish and mice without affecting the anti-neoplastic activity of doxorubicin [249]. Similarly, the endothelial cell-specific expression of B cell leukemia/lymphoma 2 related protein A1a (BCL2A1A) promotes survival in a model of allogeneic heart transplantation [250].
Finally, the mechanistic links between intrinsic apoptosis and atherosclerosis remain a matter of debate. Indeed, while Casp3 deletion favors plaque development in mouse models of atherosclerosis [251], the absence of DNA fragmentation factor subunit beta (DFFB, best known as CAD)) [252] protects mice against the disease. Likewise, while conditional deletion of Mcl1 in myeloid cells is pro-atherogenic [253], genetic or pharmacological inhibition of BCL-XL reduces atherosclerosis via a mechanism involving the depletion of platelets [254]. Moreover, the macrophage or leukocyte-specific deletion of the gene encoding BIM in mice has modest effects on plaque development, especially in the early phase of atherosclerosis [255, 256]. As the etiology of atherosclerosis involves a major inflammatory component, these apparently discrepant results may reflect (at least in part) the key role of some components of the apoptotic machinery in the control of inflammatory responses.
Renal disorders
Germline or kidney-specific deletion of Bax attenuates acute kidney damage in mice subjected to experimental renal ischemia/reperfusion [257]. A similar nephron-protection has been observed in Bid−/− mice [258], as well as in transgenic mice specifically expressing BCL-XL in the kidney [259]. Moreover, the simultaneous deletion of Bax and Bak1 in kidney proximal tubules limits tubular apoptosis and ameliorates kidney inflammation and fibrosis in a mouse model of renal fibrosis based on unilateral ureteral obstruction [260, 261]. Apoptotic caspases also appear to contribute to the etiology of renal conditions, although this may reflect cell-extrinsic effects of caspase activity. Casp3 deletion reduces microvascular rarefaction and renal fibrosis in mice subjected to experimental ischemia-reperfusion injury [262], resulting in better long-term outcomes [263]. Moreover, the lack of CASP3 increases the survival of mice with chronic kidney disease caused by a congenital mutation in cystin 1 (Cys1) [264]. In this setting, CASP3-deficient mice display increased CASP7 and decreased BCL2 expression, which is in line with recent clinical evidence of constitutive BCL2 down-regulation in patients with polycystic kidney disease [265]. Administration of broad-spectrum caspase inhibitors limits kidney damage and improves renal functionality after a variety of experimental insults to kidneys, as observed in animal models of renal ischemia [266, 267], polycystic kidney disease [268], glomerulonephritis [269], lupus nephritis [270] and diabetic renal disease [271]. Nonetheless, the specific targeting of apoptotic caspases will reveal whether this effect reflects the inhibition of intrinsic apoptosis. Indeed, these studies do not rule out the involvement of non-apoptotic RCD pathways in the etiology of acute and chronic kidney injury [272, 273]. Moreover, some of the nephron-protective effects of broad-spectrum caspase inhibitors have been linked to decreased post-RCD inflammation rather than the sole inhibition of apoptosis [266, 274]. In this context, Z-VAD-FMK aggravates (rather than ameliorates) renal dysfunction in a mouse model of cisplatin nephrotoxicity, by a mechanism involving the abrogation of cyto-protective autophagy [275]. Similarly, Z-VAD-FMK is ineffective in mouse models of osmotic nephrosis and contrast-induced acute kidney injury [276], and this may be linked to the ability of Z-VAD-FMK to inhibit CASP8 (and hence promote necroptosis). Finally, acute loss of BCL-XL in all tissues of adult mice, except for hematopoietic cells, caused severe renal tubular degeneration leading to fatal anemia due to the loss of erythropoietin production [277].
Hepatic diseases
Abundant evidence highlights pathogenic roles of apoptosis in acute liver injuries, as well as in alcohol-related and alcohol-unrelated chronic liver disorders. Hepatocytes express high levels of BID, which connects DR signaling to mitochondrial outer membrane permeabilization (MOMP) upon CASP8-dependent cleavage [278], and this complicates distinguishing between the intrinsic and extrinsic pathways. Here, we will focus on studies performed in animal models of liver injury unrelated to overt signaling engaged by the Fas cell surface death receptor (FAS; also known as CD95 or APO-1) or TNF receptor superfamily member 1A (TNFRSF1A, best known as TNF-R1), which instead will be discussed in the next section.
Distinct preclinical models of hepatic ischemia-reperfusion injury demonstrated that deletion of Bcl2l11 (leading to lack of BIM) and/or Bid as well as over-expression of BCL2 or administration of pharmacological broad-spectrum caspase inhibition mediate robust hepatoprotective effects [279,280,281,282]. A similar improvement of hepatocyte survival and liver functionality was observed in rodents specifically expressing a mutated variant of BID in the liver and subjected to warm ischemia/reperfusion injury [283]. As for other models of liver injury, BIM-deficient mice are protected against viral hepatitis [284]. Moreover, deletion of the genes encoding BIM or PUMA, but not BCL2-related ovarian killer (Bok) limits liver injury in mice exposed to the hepatotoxic agent acetaminophen [285,286,287]. Moreover, pre-treatment with Z-VAD-FMK improves the survival of mice subjected to extensive hepatectomy [288].
There is contrasting evidence on the role of BID in the etiology of liver conditions unrelated to overt FAS and TNF-R1 signaling. In a model of alcohol-related liver disease, the lack of BID confers some protection against ethanol-induced fibrosis, although mice display persisting signs of inflammation and steatosis [289]. Moreover, mice with a hepatocyte-specific deletion of Bid present reduced liver inflammation and fibrosis when subjected to a choline-deficient diet to cause non-alcoholic steatohepatitis (NASH) [290]. Also, administration of BID-targeting antisense oligonucleotides exerted significant hepatoprotective effects [291]. However, BID deficiency fails to ameliorate liver injury and fibrosis upon bile duct ligation (as a model of obstructive cholestasis and chronic liver disease) [292]. Of note, in the same experimental model, the liver-specific overexpression of MCL1 but not BCL2 protects animals from hepatic damage [293, 294], suggesting some specificity for MCL1. To add a layer of complexity, conditional deletion of Xiap in hepatocytes does not result in liver injury, steatosis, or fibrosis, possibly due to compensatory effects of other inhibitor of apoptosis protein (IAP) isoforms [295]. That said, Xiap−/− and Casp3−/− mice subjected to diet-induced hepatic steatosis and/or fibrosis, display exacerbated and attenuated liver damage, respectively [296, 297]. These effects have been linked to the modulation of the inflammatory response rather than apoptosis. Finally, genetic co-deletion of Mcl1 and transformation-related protein 53 (Trp53, best known as p53) [298], as well as conditional deletion of the genes encoding BCL-XL or MCL1, promote fibrosis and/or carcinogenesis, two common final stages of liver disease [299]. In this latter study, the additional deletion of Bak1 limited hepatotoxicity, which is in line with evidence indicating that deletion of Bid and/or Bok protects mice against experimentally induced hepatocarcinogenesis [300,301,302].
CASP2 was found to be upregulated in a mouse model of NASH and in NASH patients, and was implicated in driving lipogenesis and steatohepatitis with a mechanism involving the cleavage of the site-1-protease (S1) followed by the activation of sterol regulatory element binding proteins (SREBP) [303]. In this study, the ablation or pharmacological inhibition of CASP2 prevented diet-induced steatosis and NASH progression. Of note, CASP2 deficiency was also reported to protect mice from diet-induced obesity and metabolic syndrome [304]. Supporting the etiological contribution of caspase activation to liver disease, the administration of broad-spectrum caspase inhibitors (e.g., emricasan, VX-166) reduced liver injury, inflammation and fibrosis in mice fed a diet rich in fat or deficient in methionine and choline [305, 306]. Along similar lines, emricasan reportedly decreased portal pressure, fibrogenesis and hepatic inflammation, and preserved liver function in rodent models of chronic carbon tetrachloride (CCl4)-mediated cirrhosis or cholestasis driven by bile duct ligation [307,308,309]. Preliminary anti-inflammatory effects coupled with improved liver function have also been observed in patients with NASH-related cirrhosis treated with emricasan [39, 310]. However, follow-up, randomized clinical studies failed to observe beneficial effects of this agent on portal pressure and clinical outcome [40, 41, 311]. At least in part, these findings may reflect the complex interconnection between multiple RCD variants involved in the pathogenesis of NASH. Supporting this possibility, the administration of CASP3-specific inhibitors that abrogate both pro-apoptotic and pro-pyroptotic activities of CASP3 protected mice against acute liver injury caused by bile duct ligation [312]. Additional pharmacological and genetic studies specifically targeting intrinsic apoptosis (over other RCD pathways controlled by caspases) are needed to formally ascertain the involvement of this pathway in the etiology of hepatic disorders.
Hematological malignancies and solid cancers
The role of intrinsic apoptosis in preventing oncogenesis has been demonstrated in multiple animal models of induced hematological and solid tumors. In particular, a wide range of evidence demonstrates that over-expression of BCL2, BCL-XL or MCL-1 accelerates the onset of leukemia and lymphoma induced by over-expression of the MYC proto-oncogene, bHLH transcription factor (MYC) [313,314,315,316,317]. Accordingly, the pharmacological inhibition of anti-apoptotic BCL2 proteins is effective against MYC-driven tumors, even when they lack p53 functions [318,319,320,321]. In this context, p53 has been shown to exert multiple roles in RCD (e.g., [322,323,324]). In particular, it acts as a direct or indirect regulator of the expression of several apoptotic genes [325,326,327,328] and connects apoptosis induction and cell cycle arrest [329]. One main target of p53 is cyclin dependent kinase inhibitor 1A (CDKN1A, best known as p21). p53-induced expression of p21 leads to the activation of DREAM and RB/E2F transcriptional repressor complexes, in turn promoting cell cycle arrest by downregulating crucial cell cycle regulators such as cyclins and cyclin-dependent kinases [326, 327, 330]. However, recent finding indicates that the p53-p21-DREAM or p53-p21-RB/E2F axis can also downregulate CASP2 and CASP8-associated protein 2/FLASH (CASP8AP2), generating a feedback loop centered on p53 that limits rather than promoting the induction of apoptosis [326, 327]. Of note, when analyzing the impact of endogenous proteins, it was shown that the absence of BCL-XL but not BCL2 limits the development of lymphoma in transgenic mice expressing MYC under the IgH enhancer (Εμ-myc mice) [331, 332], thus supporting the therapeutic use of BCL-XL inhibitors against these hematological cancers. Along similar lines, MCL1 overexpression [317] or Mcl1 ablation [318, 333, 334] accelerates and suppresses MYC-driven lymphomagenesis, respectively. Lending further support to the relevance of MCL1, prevalence and onset of MYC-driven lymphoma development were reduced by Mcl1 haploinsufficiency [318, 334], or B cell-specific deletion of Mcl1 [335]. Of note, loss of one allele of Mcl1 (but not deletion of the gene encoding BCL-XL) also impairs the development of thymic lymphoma in p53-deficient mice [336], which possibly explains the limited effect of the BCL-XL, BCL2 and BCL-W inhibitor ABT-737 in these models of tumorigenesis [337]. The contribution of pro-survival BCL2 proteins in the development of AML has been demonstrated by using mice reconstituted with genetically modified bone marrow cells overexpressing MYC [338] and in human Burkitt lymphomas and diffuse large B-cell lymphomas [339]. Notably, the acute genetic removal of Mcl1 prevents the sustained survival and proliferation of AML driven by diverse oncogenic fusion proteins [340]. Accordingly, MCL-1 specific BH3 mimetic drugs, such as S63845, are able to potently kill a diverse range of lymphoid and myeloid malignant cells in culture and even in tumor transplanted mice [341]. Finally, ablation of Bcl2l2 (leading to lack of BCL-W) limits the development of MYC-mediated B cell lymphoma [342].
Numerous studies demonstrated that the development of MYC-driven lymphoma and leukemia is accelerated in mice lacking the genes encoding BAX [343], BIM [344, 345], BAD [346], BMF [346] or PUMA [347,348,349]. In particular, these studies report that loss of only a single allele of Bcl2l11 (encoding BIM) accelerates the development of lymphoma and this effect can be reversed following full ablation of Bcl2l1 (leading to lack of BCL-XL) [345]. In this context, the presence of all prosurvival BCL2 proteins is shown to limit the impact of BIM in Eμ-Myc transgenic mice [350]. Instead, the combined ablation of the genes encoding BIM and p53 or PUMA and p53 accelerates MYC-driven lymphomagenesis [351]. This is in line with the evidence that loss of the genes encoding BAX or BIM augmented lymphomagenesis in p53-deficient mice [352, 353]. Of note, PUMA seems to exert a strong tumor-suppressive role in hematological cancers, as shown by the evidence that Bbc3 deletion accelerates the development of MYC-driven B-cell lymphomas and that Eμ-Myc lymphomas developing in PUMA-proficient mice display downregulated expression of PUMA [348, 349, 354]. On the contrary, the loss of the gene encoding NOXA does not accelerate MYC-driven lymphomagenesis, and the role of BIK in this murine lymphoma model is debated [348, 355]. Along similar lines, while CASP2 suppresses MYC-induced lymphomagenesis in mice [356], the tumor suppressive role of apoptosome components (Box 1) is questioned, as shown in lethally irradiated mice reconstituted with Eμ-Myc transgenic APAF1-deficient or CASP9-deficient fetal liver cells which showed no difference in the incidence of lymphoma compared to their wild-type counterparts [357]. This is consistent with the notion that APAF1 and caspase-9 function downstream of the commitment to cell death (MOMP) and therefore do not act as tumor suppressors [15].
Concerning other experimental animal models of hematological malignancies, the absence of PUMA (due to ablation of Bbc3) abrogated the development of both myelodysplasia, as shown in transgenic mice expressing a nucleoporin 98 (NUP98)-homeobox D13 (HOXD13) fusion protein [358], and thymic T cell lymphoma induced by gamma radiation [359, 360]. The explanation for these surprising findings is based on the fact that the absence of PUMA prevents the extensive death of hematopoietic cells caused by gamma radiation, which causes mobilization and extensive proliferation of hematopoietic stem and progenitor cells, resulting in elevated replication stress and genetic instability and lymphomagenesis. These observations show that inhibition of apoptosis does not only promote the development of hematological malignancies, but in certain conditions can do the exact opposite and prevent lymphoma development. The absence of NOXA, augments the development of chronic lymphocytic leukemia in T cell lymphoma breakpoint 1 (TCL1) transgenic mice [361] and accelerates the development of thymic T lymphoma induced by gamma radiation [359]. Moreover, conditional deletion of Bcl2l11 in B cells (leading to the absence of BIM) accelerates the development of mantle cell lymphoma in mice driven by cyclin D1 (CCND1) overexpression [362]. Overexpression of MCL1 and/or BCL2 promotes the development of acute myeloid leukemia driven by lysine (K)-specific methyltransferase 2A (KMT2A, best known as MLL) fusion proteins [340, 363] and plasmacytoma driven by ABL proto-oncogene 1, non-receptor tyrosine kinase (ABL1) [364]. Conversely, the loss of one Mcl1 allele suppresses the development of T cell lymphoma, as shown in models based on sequential low-dose irradiation or the expression of a transgene encoding an IL2 inducible T cell kinase (ITK)-spleen tyrosine kinase (SYK) fusion protein [365]. Finally, the absence of CASP2 accelerates lymphomagenesis in ataxia telangiectasia mutated (ATM)-deficient mice [366], but this may be due to the loss of the function of CASP2 in mitotic cell division [367]. Lending support to the role of intrinsic apoptosis in hematologic malignancies, the BCL2 inhibitor venetoclax has entered clinical practice for the treatment of CLL as single agent or more effectively in combination with other therapeutic agents [31, 35,36,37]. Combinatorial regimens of BCL2 inhibition with epigenetic modulation have entered center stage in certain settings of AML [38, 368]. However, mechanisms of resistance of CLL and AML to venetoclax related to defects in p53 and the apoptotic network or deregulated energy metabolism have been described [369,370,371,372]. Venetoclax-based regimens also display effectiveness in patients with high-risk myelodysplastic syndromes [373], suggesting a potential application of venetoclax to other hematological cancers [374, 375].
Significant work demonstrated a tumor suppressor role of the intrinsic apoptotic pathway in many cancers. For example, BCL2 overexpression accelerates the development of MYC-induced mammary tumorigenesis [376]. A similar acceleration of tumor development has been described for the loss of genes encoding BAX, BIM, CASP2 or PUMA in distinct models of breast cancer induced by expression or overexpression of C3(1)/SV40 T-antigen, MYC, or erb-b2 receptor tyrosine kinase 2 (ERBB2, best known as HER2) [377,378,379,380]. At odds with these results, BCL2 overexpression in the mammary gland suppresses the development of breast tumors driven by the administration of dimethylbenz(a)anthracene [381]. This latter finding may be explained in a similar way as was mentioned for the suppression of radiation-induced thymic T cell lymphoma development by over-expression of BCL-2 or loss of PUMA (see above). Conditional deletion of the genes encoding BCL2 or BCL-XL in intestinal epithelial cells delays the development of colorectal cancer driven by inflammation [382, 383], which is in line with the evidence that the absence of PUMA (due to Bbc3 deletion) exacerbates colorectal tumorigenesis as shown in a mouse model of intestinal oncogenesis driven by colitis or APC, WNT signaling pathway regulator (APC) [384]. Interestingly, doxorubicin-induced intestinal cytotoxicity requires PUMA but not BIM, whereas the reverse is true for MYC-driven apoptosis in the gut, indicative of differential roles for different BH3-only proteins in this tissue [385]. Intriguingly, treatment with BCL-XL, but not BCL2-targeting BH3 mimetics is sufficient to prevent intestinal tumorigenesis, suggesting that BCL-XL is the crucial mediator of protection of early neoplastic cells in this model [386]. In agreement, earlier work showed a pronounced BCL-XL dependency of cell lines derived from both colorectal and non-small cell lung cancers [387, 388]. Moreover, a tumor suppressive effect has been ascribed to BAX and CASP2 in murine models of brain [389, 390] and lung [391] oncogenesis, respectively. In line with this evidence, pharmacologic/genetic inhibition of MCL1 delayed tumor development in a mouse model of mutant KRAS-driven adenoma/adenocarcinoma [392]. In the same model, tumor progression was promoted by the ablation of pro-apoptotic Bok [393]. Of note, there is evidence of a certain tissue-specificity in the epigenetic regulation of Bcl2 and Mcl1, such as the epigenetic mechanism centered on the deubiquitinase BRCA1 associated protein 1 (BAP1) [394], a tumor suppressor that is frequently mutated in some cancers [395] and has been associated with tumor aggressiveness and therapy resistance [396, 397]. Finally, age-related differences in the expression of pro-apoptotic members of the BCL2 family have been linked to the increased sensitivity of neonatal/childhood tissues, relative to adult counterparts, to chemotherapy and radiotherapy. This was causally linked to the MYC-dependent expression of genes encoding BAX, BID and BIM, both in mice and humans [133].
Cancer-specific roles have been attributed to particular BCL2 protein family members. For example, deletion of Bax accelerates the development of MYC-induced pancreatic tumors [398], which was not seen with ablation of Bak1 or Casp3 [398, 399], but was achieved by BCL-XL overexpression [314, 400]. Likewise, BOK seems to be crucial in hepatocarcinogenesis, as demonstrated in a mouse model of diethylnitrosamine-induced liver cancer which was accelerated on a Bok−/− genetic background [300]. Using the same mouse model, accelerated hepatic carcinogenesis has also been demonstrated for the deletion of the genes encoding PUMA or CASP2 [401, 402]. Conversely, overexpression of BCL2 was shown to limit transforming growth factor alpha (TGFA)-driven hepatic tumorigenesis [403, 404], perhaps because the death of some cells in the liver causes massive mobilization and proliferation of progenitor cells, leading to acquisition of oncogenic lesions that drive tumorigenesis in a manner similar to radiation-induced thymic lymphoma development (see above). Finally, the transgenic overexpression of BCL-XL (but not BCL2) and the keratinocyte-specific deletion of Bcl2l1 (leading to lack of BCL-XL) accelerates or limits, respectively, carcinogen- and/or ultraviolet B (UVB)-induced skin tumorigenesis [405,406,407,408]. It will be important to investigate and better understand why in specific settings inhibition of apoptotic cell death promotes tumorigenesis whereas it inhibits tumorigenesis in others.
Autoimmune and inflammatory diseases
There is substantial evidence linking intrinsic apoptosis to the development and progression of autoimmune diseases. However, the interpretation of these findings should take into consideration the crosstalk between the apoptotic and inflammatory pathways and the fact that apoptotic caspases accelerate cell death as they regulate its immunological manifestations.
The first evidence that defects in the intrinsic apoptosis pathway can cause the development of autoimmune disease was reported when over-expression of BCL-2 in B lymphocytes [409] or loss of BIM in all tissues [410] was shown to cause a fatal systemic lupus erythematosus (SLE)-like disease. Consistent with a critical role for the intrinsic apoptotic pathway in preventing autoimmune disease, the combined loss of the genes encoding BAX and BAK1 in hematopoietic cells, achieved by transplantation of lethally irradiated wild-type mice with hematopoietic stem/progenitor cells from the livers of E14.5 Bax−/−Bak1−/− embryos also causes a fatal SLE-like disease [411]. In mouse models of rheumatoid arthritis, ablation of the genes encoding BIM, BID or BAD, but not the loss of Bax and Bak1, accelerated the emergence and increased the duration and severity of this disorder [412,413,414]. Consistent with these findings, administration of a BIM mimetic suppressed inflammatory arthritis in mice [415]. Mice deficient for BAX as well as transgenic mice expressing XIAP display increased severity of autoimmune encephalomyelitis induced by immunization with myelin oligodendrocyte glycoprotein (MOG) [416, 417]. Similar results have been obtained in mouse models of autoimmune encephalomyelitis genetically engineered for the hematopoietic cell-specific deletion of Bcl2l11 (leading to BIM deficiency), or the neuron-specific overexpression of BCL2 [418, 419]. Consistent with the notion that inhibition of apoptosis can promote the development of auto-immune disease, inhibition of BCL2, BCL-XL and BCL-W using the BH3 mimetic ABT-263 substantially reduced pathology in several mouse models of autoimmune disease, including scleroderma [420]. In apparent contrast with these results, studies using models of type 1 (autoimmune) or type 2 (non-autoimmune) diabetes revealed that deletion of Bax alone or combined loss of Bax and Bak1 [421, 422], deletion of the gene encoding BIM, alone or together with the gene encoding PUMA [418, 423,424,425] as well as the loss of BMF [426], protect pancreatic β cells from autoimmune destruction. Moreover, the absence of BIM prevents the emergence of type 1 diabetes in non-obese diabetic (NOD) mice [418, 423], while ablation of Trp53 in pancreatic β cells failed to halt cell death in multiple experimental models of diabetes [427].
Based on the studies described above, inhibiting or deleting pro-apoptotic proteins or genes can have conflicting effects on autoimmune disease progression. This may depend on the cell type in which the major effect on apoptosis occurs, e.g., the immune cells or their targets. Inhibiting cell death in the target cells would indeed provide protection and may improve disease outcome, whereas inhibiting cell death in the immune cell may lead to an accumulation of immune cells and aggravation of the autoimmune disease. The distinction could be explored by studying tissue-specific deletion of apoptosis regulator genes.
In this context, there is evidence that inflammatory and autoimmune disorders may derive from increased survival of specific immune cell population. For instance, elevated levels of cytokines such as colony stimulating factor 2 (CSF2, best known as GM-CSF), interleukin 3 (IL3) and IL5 in immune disorders have been associated with prolonged survival of neutrophils, eosinophils or basophils with a mechanism involving the upregulation of anti-apoptotic proteins MCL1, BCL-XL and baculoviral IAP repeat containing 2 (BIRC2, best known as cIAP2) [428,429,430,431,432,433,434,435]. Apoptosis also plays a relevant role in some hemopathies with inflammatory features, including beta thalassemia [436], Diamond-Blackfand anemia [437], and in the Cohen syndrome neutropenia [438]. BIM, BID and BAD have all been shown to influence survival in mouse models of septic shock, as their targeting confer protective effects from tissue damage of multiple organs [439,440,441], as well as in patients with severe sepsis [442]. On the contrary, the role of apoptotic caspases in septic shock is contentious [54, 73, 443, 444]. The precise impact of apoptosis in widespread inflammation during sepsis requires further investigation.
Concerning other inflammatory diseases, while broad-spectrum caspase inhibition reportedly protected rats against severe acute pancreatitis [445], activation of intrinsic apoptosis appears to attenuate the severity of this disease by limiting inflammation, as shown in vivo in a pancreatitis mouse model lacking XIAP [446]. These data reinforce the notion that inhibiting (apoptotic) cell death may exacerbate unwarranted inflammatory reactions that contribute to the pathology of various autoimmune and inflammatory disorders. In line with this notion, chronic colitis driven by dextran sulfate sodium in mice manifests with increased (rather than decreased) severity in BID- or BIM-deficient hosts as compared to their wild-type littermates, at least in part owing to immune dysregulation [447, 448]. Similarly, inhibition of BCL2 and/or BCL-XL reduces inflammation and ameliorates experimental colitis [449, 450], an effect that was abrogated by concomitant deletion of the gene encoding BIM [450]. PUMA-deficient mice display reduced levels of apoptosis amongst intestinal epithelial cells but not reduced inflammation in an experimental model of colitis [451]. Corroborating the specific relevance of PUMA for intestinal homeostasis, mice deficient for PUMA but not Bax−/−Bak1−/− mice were protected against the gastrointestinal side effects of radiotherapy, at least in part due to increased survival of intestinal stem/progenitor cells [452, 453]. Moreover, the absence of PUMA conferred protection to intestinal epithelial cells in mouse models of hypertensive gastropathy [454], ulcerative colitis (UC) [455] and intestinal ischemia/reperfusion [456]. In the latter model, transgenic BCL2 expression limited intestinal epithelial cell death [457]. On the other hand, defects in XIAP cause X-linked lymphoproliferative syndrome type 2, with one-third of these patients suffering from severe and therapy-refractory inflammatory bowel disease [458,459,460,461]. Absence of XIAP also results in enhanced TNF production and TNF-R1/TNF-R2 targeting of TLR5-expressing Paneth cells and dendritic cells (DCs), leading to ileitis and dysbiosis [462]. In this context, it is interesting to note that CASP3- or CASP7-deficient mice display an altered gut microbiome [463], which may play a hitherto unexplored role in multiple autoimmune and inflammatory disorders beyond intestinal conditions. However, it has recently been found that under steady state conditions the absence of CASP3 and CASP7 in the intestinal epithelial cells apparently neither affects the microbiome nor causes spontaneous inflammation, suggesting that apoptosis may be dispensable for intestinal epithelium turnover and homeostasis at baseline [464].
Infectious diseases
Activation of RCD constitutes a protective mechanism against many microbial infections by eliminating infected cells and potentiating pathogen-targeting immune responses. Accordingly, both viruses and bacteria have developed multiple strategies to overcome or disable host intrinsic apoptosis, thus improving survival of both host cells and the infectious organisms [465, 466]. Mice with loss of one BCL-XL-coding allele displayed reduced pathology and had improved survival rates when challenged with Japanese encephalitis virus (JEV), as compared with wild-type mice. This was attributed to compromised viral propagation within JEV-infected cells succumbing to intrinsic apoptosis [467]. There is also evidence of a contribution of BAX and BAK1 to the response to murine cytomegalovirus (MCMV) infection. In particular, the MCMV genome encodes inhibitors of BAK1 (m41.1 protein) and BAX (m38.5 protein) that promote viral replication by inhibiting the induction of intrinsic apoptosis in infected cells [468, 469]. Supporting the requirement of the inhibition of intrinsic apoptosis for optimal in vivo MCMV dissemination, the titers of m41.1-deficient viruses were higher in salivary glands and other organs in Bak1−/− mice as compared to wild-type animals [468]. Intrinsic apoptosis also protects against bacterial infections, as demonstrated by the lethal course of disease in Bbc3−/− mice (which lack PUMA) after Streptococcus pneumoniae infection [470]. Such an effect has been attributed to insufficient immune-mediated bacterial clearance because of an increased neutrophil lifespan in the absence of PUMA-mediated apoptosis.
However, in other contexts, excessive activation of the intrinsic apoptosis pathway has been reported to drive, rather than prevent, microbial disease pathogenesis and lethality. For example, loss of Xiap increased the susceptibility of mice to Shigella infection, manifesting with coalescing necrotic areas and a high bacterial burden in the liver, an effect that was associated with an inefficient immune-mediated resolution of the bacterial infection [471]. Of note, at least part of this effect may be due to the requirement for XIAP to activate NOD signaling, rather than its ability to inhibit caspases [459, 471, 472]. Moreover, mice lacking the genes encoding BIM and NOXA (i.e., Bcl2l11−/−Pmaip1−/− mice) display pronounced resistance to challenge with high doses of Listeria monocytogenes, as shown by a decreased bacterial burden and reduced apoptosis induction in the spleen [473]. The overexpression of BCL2 in the hematopoietic compartment increase the survival of mice infected with Ebola virus [474], while deletion of Bok promote resistance of lung epithelial cells to apoptosis induced by SARS-CoV-2 virus membrane (M) protein [475]. Intriguingly, this latter study showed that the SARS-CoV-2 M protein activate BOK to trigger apoptosis in the absence of BAX and BAK1 [475]. In another example, conditional deletion of Casp3 in the murine intestinal epithelium conferred protection from pathogenic Salmonella enterica, and this was attributed to a reduction in cell death-induced nutrients that are critical for sustaining bacterial growth [476]. Finally, Casp3−/− mice subjected to intracranial inoculation of reovirus type 3 (strain Dearing) displayed limited injuries in the central nervous system (CNS) and extended survival compared to wild-type mice [477]. As discussed above, the interpretation of the infection phenotypes observed in CASP3-, CASP7- and/or CASP9-deficient mice requires particular caution because of the crucial roles of these caspases in modulating immune and inflammatory responses [242,243,244]. That said, there is evidence for a role of specific regulators of apoptosis in the host response to infection with human herpes simplex virus 1 (HSV-1). On the one hand, a significant accumulation in total leukocyte and CD8+ T cells was observed in mice deficient for BIM and PUMA upon infection with HSV-1 [478], which is in line with a role of these BH3-only proteins in controlling the survival of lymphoid and myeloid cells [410, 479, 480]. On the other hand, mice deficient for NOXA, BAD or BID were reported to mount a normal CD8+ T cell immune response to HSV-1 infection [478]. Some of these contradictory results may arise from the divergent effects of inhibition or promotion of apoptosis on immune cells versus other cell types affected by the infectious disease, a distinction that cannot be addressed using mice in which apoptotic regulators have been deleted in the germline. In this context, it is noteworthy to note that the myeloid cell-specific deletion of the gene encoding BCL-XL or its inhibition using BH3 mimetic drugs massively reduced bacterial burden in the lung and extended the survival of mice infected with Legionella [481]. This indicates that BH3 mimetic drugs might be effective for the treatment of intracellular bacterial infections.
Other diseases
Pro-apoptotic BCL2 proteins and caspases have also been implicated in disorders affecting other tissues/organs, such as skeletal muscle and lungs. For instance, the conditional ablation of Bax and Bak1 protected mouse skeletal muscles against pressure-induced injury [482]. Similar results have been obtained in rats receiving Z-VAD-FMK after being subjected to muscular compression or blunt injury [483, 484]. Moreover, deletion of Casp3 or CASP3 inhibition with Ac-DEVD-CHO limited muscular damage and atrophy in experimental models of plaster-mediated immobilization [485, 486]. In mouse models of catabolic disorders, muscle wasting due to protein degradation was decreased by lentiviral expression of XIAP [487, 488], although whether this effect reflects the inhibition of intrinsic apoptosis needs further confirmation. Finally, Casp3−/− mice were protected against denervation-induced muscular atrophy [489], while expression of a dominant-negative variant of CASP9 improved the neuromuscular activity in a transgenic mouse model of slow-channel syndrome [490].
In a mouse model of oxidant-induced lung injury, the tissue-specific ablation of Bax and Bak1 but not that of the genes encoding BID, BIM, NOXA or PUMA protected lung epithelial cells from degeneration [491]. Among the anti-apoptotic BCL2 proteins, BCL2 related protein A1 (BCL2A1, best known as A1) seems to exert a crucial role in this setting, as Bcl2a1 deletion aggravated lung injury in mice subjected to hyperoxia [492], while lung-specific overexpression of BCL2 did not confer protection to mice exposed to excessive oxygen supply [493]. That said, no critical cytoprotective effect of A1 was seen in acute lung inflammation and peritonitis [494]. Intrinsic apoptosis has also been reported to be involved in pulmonary fibrosis [495]. Bid−/− mice display decreased levels of pulmonary fibrosis after intra-tracheal bleomycin administration than their wild-type counterparts [496]. In apparent contradiction, in the same model of fibrotic pulmonary damage, a similar degree of protection was reported in mice lacking Bcl2 [497] or in animals treated with inhibitors of BCL2 [497] or caspases [498, 499]. Along similar lines, ablation of Bid limited acute lung injury in mice induced by exposure to lipopolysaccharide (LPS) [500]. Moreover, CASP3 depletion using short-hairpin RNAs (shRNAs) protected the lungs of mice subjected to pulmonary ischemia/reperfusion [501], a protection further strengthened when necroptosis was concomitantly also suppressed [502]. BCL2 overexpression or caspase inhibition protected rodents subjected to lung transplantation [503, 504]. This is in line with the notion that delivery of the caspase inhibitor Z-VAD-FMK to rodents ameliorated lung injury developing as a consequence of severe acute pancreatitis or LPS administration [505, 506] but not as a result of pneumovirus infection [507]. In the latter case, lung damage was exacerbated by Z-VAD-FMK, perhaps due to increased inflammation downstream of necroptotic RCD [507].
The studies summarized above illustrate that components of the intrinsic apoptosis pathway can be part of the pathogenic mechanism of disease, and, in certain cases, this may offer the opportunity for therapeutic intervention. It is important to note that in many pathogenic processes intrinsic apoptotic cell death is the endpoint, and simply inhibiting it will not be curative. If the cells continue being exposed to the initiating insult, they will likely undergo less regulated forms of cell death. However, inhibiting the intrinsic apoptotic cell death may buy time to control the factors that are damaging the cells in first place. Ischemia and hypoxia, in cases where the ensuing cell death has a substantial intrinsic apoptotic component, are examples. If cells in the ischemic region were kept alive until adequate circulation was restored, therapeutic benefits might be achieved. Other examples include metabolic disorders, which may be amenable to correction, and traumatic injury, where healing might be supported by inhibiting apoptosis. It would be worth concentrating on inhibiting intrinsic apoptotic cell death in conditions where the initiating tissue insults can be (at least partially) reversed. In contrast, failure to undergo intrinsic apoptosis is the initial pathogenic step or a contributing factor in certain malignancies. Here, the induction of apoptosis, for example by using BH3 mimetic drugs [33, 34], directly targets pathogenesis.
Extrinsic apoptosis in disease
The molecular apparatus for extrinsic apoptosis is described in Box 5 and illustrated in Fig. 4. Unlike the intrinsic apoptotic pathway, DR-induced apoptosis is not required for embryonic or fetal development but plays a critical role in adult tissue homeostasis, as detailed in Box 6 and Box 7. Of note, various components of the extrinsic pathway of apoptosis are involved in the etiology of multiple human disorders, although (1) with a considerable degree of context-dependency, and (2) with an effect not necessarily linked to the activation of apoptosis but often due to the role of DR signaling in necroptosis and inflammation, as outlined below.

Extrinsic apoptosis is initiated by the binding of FASL to FAS or TRAIL to TRAIL-R1 or TRAIL-R2, which promotes the assembly, on the cytoplasmic tail of these death receptors, of a platform known as the DISC. Extrinsic apoptosis is also triggered by the binding of TNF to TNF-R1, which promotes the assembly of the Complex II. The DISC comprises FADD, c-FLIPs and pro-CASP8. Complex II is a platform consisting of FADD and pro-CASP8 in association with either TRADD (complex IIa) or RIPK1 (complex IIb). The assembly of these complexes promotes the activation of CASP8, which mediates CASP3 and CASP7 activation either directly, by catalyzing the proteolytic activation of CASP3 and CASP7 (in type I cells) or indirectly, via the proteolytic activation of the BH3-only protein BID and outer membrane permeabilization (MOMP) (in type II cells). At least in some cells, extrinsic apoptosis can also be induced by dependence receptors like DCC, NTRK3, PTCH1, or UNC5A-D, which are activated by decreased concentration of the related ligand, as illustrated in the figure. However, the role of this pathway in normal physiology and disease is not yet established.
Neurological diseases
Although numerous studies investigated FAS and TNF-R1 signaling in the pathogenesis of multiple neurological diseases, the precise role of extrinsic apoptosis remains unclear (Fig. 5). Loss-of-function mutations of Fas ligand (TNF superfamily, member 6) (Fasl) as well as Fas silencing prevented moto-neuron loss in mouse models of ALS driven by defect in superoxide dismutase 1, soluble (SOD1) [508, 509]. Conversely, the lack of TNF did not affect motor neuron loss and mouse survival in this model [510], while signaling via TNF receptor superfamily member 1B (TNFRSF1B, best known as TNF-R2) appeared to mediate neuroprotective effects [511]. As an additional layer of complexity, TNF mediates neuroprotective functions in wobbler mice - another mouse model of ALS that carries a point mutation in VPS54 GARP complex subunit (Vps54), at least in part by promoting the upregulation of ADAM metallopeptidase domain 8 (ADAM8) [512]. CASP8 has not yet been implicated in the pathogenesis of ALS, and non-apoptotic forms of FAS-driven RCD may play a predominant role in this context. For example, FAS stimulation reportedly triggered the demise of motoneurons in mouse models of ALS by aggravating endoplasmic reticulum stress [513]. Similarly, cleavage of BID by CASP1 (and not CASP8) appears to contribute to neurodegeneration in transgenic mice expressing a mutant form of human SOD1 [514]. However, the precise contributions of endoplasmic reticulum stress and CASP1 in ALS and other motoneuron disorders remain to be elucidated.

Death receptor-induced apoptosis is directly or indirectly involved in the pathogenesis of multiple neurological disorders, including neurodegenerative diseases, in brain damage due to traumatic injury or neurotoxicity as well as in neuromuscular and retinal disorders.
The ability of TNF-R1 signaling to influence neurodegenerative conditions involves not only the induction of extrinsic apoptosis but also the activation of an inflammatory response. In distinct murine models of AD, deletion of Tnf, modification of its untranslated region (UTR) as well as pharmacological TNF inhibition reduced plaque formation, resulting in attenuated neurological deficits [515,516,517,518,519,520,521,522]. Mechanistic studies in mice and monkeys revealed that TNF-R1 activation stimulates the protein activator of interferon-induced protein kinase EIF2AK2 (PRKRA) network [523], which is linked to PD in humans [524]. Moreover, TNF-R1 signaling has been shown to favor microglial reactivity during neurodegeneration, culminating in neuronal loss [525]. Amelioration of disease was seen in mouse models of AD upon genetic or pharmacological inhibition of TNF-R1 [526, 527]. AD-associated neuroinflammation seems to depend on TNF-induced necroptosis rather than extrinsic apoptosis [528, 529]. Unexpectedly, AD pathogenesis was shown to be enhanced in mice bearing a co-deletion of Tnfrsf1a and Tnfrsf1b [530], a phenotype that appears to impinge on a complex network of mutual interactions between TNF-R1 and TNF-R2 signaling [531]. Such a network may also contribute to PD pathogenesis. Genetic ablation of Tnf or Tnfrsf1a plus Tnfrsf1b (leading to the lack of both TNF receptors), as well as pharmacological inhibition of TNF, were reported to protect dopaminergic neurons in murine models of PD following the administration of 1-metil 4-phenyl 1,2,3,6-tetraidro-piridina (MPTP) or 6-hydroxydopamine [532,533,534,535]. Notably, in the aforementioned experimental settings, TNF is thought to induce neuronal death in vivo by promoting microglia reactivity [536] with a complex interaction between TNF-R1 and TNF-R2 signaling [537]. Clinical evidence from AD patients subjected to perispinal administration of the TNF blockers infliximab or etanercept suggests that the inhibition of TNF can ameliorate AD [538, 539]. In contrast, a dominant-negative variant of TNF failed to protect mice against neuronal degeneration in a model of HD [540], suggesting that this approach may not be viable in patients with HD.
TRAIL/TRAIL-R signaling has also been implicated in the onset and progression of AD [541, 542]. Specifically, in a mouse model of AD, neutralization of TNF superfamily member 10 (TNFSF10, best known as TRAIL) with a monoclonal antibody resulted in decreased neuroinflammation and a reduction in cognitive defects [541]. However, these findings were not extensively validated. Similarly, the impact of FASL-FAS signaling on neurodegenerative conditions is debated. Indeed, lpr/lpr mice, which lack FAS [543] and to a lesser extent gld/gld mice, which lack FASL [543], are particularly susceptible to neuronal degeneration driven by MPTP [544]. However, contrasting results have been obtained in another study involving FAS-deficient mice treated with MPTP [545, 546]. In this context, FAS-associated factor 1 (Faf1, a FAS binding protein that can initiate or enhance apoptosis) was found to be increased in midbrain in murine models of PD [547]. Moreover, a reduction in Faf1 expression limited MPTP-induced dopaminergic cell loss [548]. Such an apparent discrepancy in results may originate from the pleiotropic role of FAS in apoptosis and inflammation and other pro-survival/regenerative signals.
CASP8 activation has been detected in the brain of both AD [549] and HD [550] patients as well as in dopaminergic neurons of MPTP-treated mice and PD patients, a setting in which BID cleavage has also been documented [119]. This is in line with the ability of the broad-spectrum caspase inhibitor Q-VD-OPH to inhibit BID cleavage and mediate neuroprotection in MPTP-treated mice and rats [551]. Of note, CASP8 was also reported to promote microglia reactivity potentially leading to neuronal loss [552,553,554]. In this context, genetic loss or pharmacological inhibition of CASP8 attenuated neurotoxicity by reducing microglial reactivity, thus extending survival of neurons, at least in part by stimulating the necroptotic death of activated microglial cells [552,553,554]. Consistent with this notion, Casp8 deletion in myeloid cells protected mice from MPTP-mediated neurotoxicity [555], suggesting that CASP8 inhibitors may be harnessed for the treatment of neurodegenerative conditions. Corroborating this idea, a pharmacological inhibitor of TNF-R1-associated death domain protein (TRADD) protected mice from disease in a model of AD-like proteinopathy driven by mutant tau [556]. However, pharmacological inhibition of CASP8 only partially prevented neuronal alterations in other models of AD [108], and even exacerbated dopaminergic neuronal necrosis in mice developing PD upon MPTP administration [557]. Moreover, rare CASP8 loss-of-function variants have been associated with AD in a large cohort of patients [558]. Thus, the precise contribution of CASP8 signaling to neurodegenerative disorders and whether this relates to its function in driving extrinsic apoptosis, inhibiting necroptosis or promoting inflammatory cytokine production remains to be formally defined. Concerning dependence receptors, Netrin 1 (NTN1) upregulation was shown to confer neuroprotection in murine models of PD, suggesting a potential role of dependence receptors in neurodegenerative disease [559].
DR signaling has also been shown to contribute to neuronal death and inflammation in preclinical models of CNS trauma. In a compression model of spinal cord injury, mice with loss of FAS (i.e., lpr/lpr mice) as well as mice treated with FASL blockers displayed reduced post-traumatic neuronal degeneration and inflammation coupled to considerable functional improvement [560,561,562]. This beneficial effect also involved reduced engagement of the intrinsic apoptosis pathway [563]. Myeloid cell-specific deletion of Fasl promoted neuronal regeneration and functional recovery in mice subjected to spinal cord injury [564]. A similar functional improvement after spinal injury was observed in mice with conditional deletion of Tnf in macrophages and neutrophils, but not in microglia [565]. Moreover, neuroprotection and limited neuroinflammation have been documented in lpr/ lpr mice subjected to traumatic brain injury [566], as well as in mice subjected to experimental spondylotic myelopathy and exposed to FASL-neutralizing antibodies [567]. Studies on mice with loss of Fas and Tnfrsf1a revealed at least some redundancy between FAS and TNF-R1 signaling in the context of experimental brain trauma [568,569,570,571,572]. Furthermore, TNF inhibition reduced damage in mice or rats experiencing spinal cord injury [573,574,575], and also reduced the appearance of signs of autonomic dysreflexia, a cardiovascular disease associated with high-level spinal cord injury [573, 576]. Interestingly, some of these studies point to a neuroprotective function for TNF-R2 [568, 570, 572], which is in line with at least some results from models of ALS [511, 531]. Moreover, several studies question a purely detrimental effect of TNF signaling in these experimental settings [577,578,579,580]. In particular, TNF was reported to support, at least in part, regeneration and long-term functional recovery in mice exposed to traumatic brain injury [578,579,580]. Conversely, TRAIL neutralization stands out as a promising strategy to promote neuronal regeneration and functional recovery based on mice with spinal cord injuries [581, 582]. In this context, injured neurons seem to undergo Fas-associated via death domain (FADD)- and CASP8-dependent RCD [583]. Accordingly, Casp8 deletion or transgenic expression of a FADD inhibitor (the glycoprotein P45) protected mice after spinal cord injury [584, 585]. Similarly, transgenic expression of a dominant negative mutant of FADD (FADD-DN) limited motoneuron loss in mice undergoing axotomy [586].
Components of the molecular apparatus for the extrinsic pathway are associated with disorders of the visual system, again in the context of both exacerbated cell death and inflammation. Thus, in mouse and rat models of optic nerve injury, deletion of Tnfrsf1a (encoding TNF-R1) or inhibition of CASP8 with Z-IETD-FMK inhibited the degeneration of retinal ganglion cells [587, 588]. Moreover, the absence of TNF-R1 (but not the absence of TNF-R2) attenuated neurodegeneration in a mouse model of retinal ischemia, despite neuronal survival not being improved [589]. Along similar lines, deletion of Tnf [590] as well as inhibition of FAS [591] or TNF [592, 593] protected mice against retinal ganglion cell death in a model of glaucoma. Similar neuroprotective effects were documented for the conditional deletion of Casp8 in astrocytes or intra-ocular Z-IETD-FMK administration [594]. In this context, the conditional inducible ablation of Casp8 from endothelial cells reduced postnatal retinal angiogenesis and pathological neovascularization in a mouse model of oxygen-induced retinopathy [595] (note that ablation of Casp8 in endothelial cells is embryonically lethal [596]; see Box 7). Moreover, CASP8 inhibition could prevent experimental neovascularization of the cornea [597]. Finally, TRAIL neutralization protected the retinal tissue from damage associated with AD in a mouse model [598].
Experimental models of ischemic stroke and hemorrhage revealed a role of DR signaling in the pathophysiology of brain damage. In models of focal ischemia induced by middle cerebral artery occlusion, lpr/lpr as well as gld/gld mice (deficient for FAS or FASL, respectively) displayed decreased infarct size and neuroinflammation [599,600,601]. Robust neuroprotection was also observed in lpr/lpr mice subjected to neonatal hypoxia-ischemia [602], as well as in lpr/lpr and gld/gld mice subjected to hyperoxia [603]. Accordingly, inhibition of FAS or FASL exerted neuroprotective effects in an experimental murine model of stroke [604, 605]. Likewise, TRAIL neutralization limited brain injury in rats and mice subjected to middle cerebral artery occlusion [601, 606] or transient ischemia-reperfusion [607]. Moreover, despite some contention in this respect [608,609,610,611], abrogation of TNF/TNF-R1 signaling by genetic or pharmacological means prevented brain injury in rodent models of intracerebral hemorrhage [612] and focal cerebral ischemia [613,614,615,616,617,618,619,620,621]. Further corroborating a pathogenic role of DR signaling, transgene-driven expression of dominant-negative CASP8 mutant and of FADD-like apoptosis regulator (CFLAR; best known as c-FLIP) attenuated brain damage after middle cerebral artery occlusion [622, 623]. This is in line with the ability of CASP8 to drive BID activation upon focal cerebral ischemia [196], as well as with the neuroprotective effects afforded by pharmacological CASP8 inhibitors seen in mice experiencing subarachnoid hemorrhage [624] or mice and rats subjected to focal cerebral ischemia [625, 626]. Importantly, FADD and CASP8 expression and/or activation have also been associated with ischemic stroke in humans [627, 628].
Perhaps surprisingly, TNF appears to protect mice against experimental seizures, not only through the engagement of TNF-R2 but also through TNF-R1 signaling [611, 629,630,631,632,633,634] and consequent modulation of NF-κB [635, 636]. Conversely, lpr/lpr mice [637], mice with neuron-specific deletion of the gene encoding TNF-R1 [638] as well as mice and rats treated with Z-IETD-FMK [585, 639, 640] display reduced sensitivity to experimental seizures, pointing to a detrimental role for apoptotic DR signaling in this condition. The precise mechanisms through which TNF-R1 signaling promotes anti-apoptotic and anti-inflammatory effects in the context of excitotoxic insults remain unclear.
Cardiovascular disorders
Data from preclinical models of ischemic and non-ischemic conditions indicate the involvement of FASL, TRAIL and TNF in the onset and progression of myocardial infarction with reperfusion and other heart diseases. In particular, both lpr/lpr mice (lacking FAS), as well as hearts isolated from these animals, displayed reduced cardiomyocyte death and infarct area upon experimental ischemia-reperfusion [641, 642]. Nonetheless, no protection against ischemia-reperfusion was found in hearts from Fas−/− or Fasl−/− mice [643]. However, supporting the therapeutic potential of the inhibition of DR signaling for the management of myocardial infarction, FASL-neutralizing antibodies conferred cardioprotection, limited inflammation, and improved cardiac function in mice experiencing cardiac ischemia-reperfusion [644,645,646]. Likewise, TRAIL blockade protected monkeys, pigs, and rats against experimental infarction by increasing cardiomyocyte survival and reducing inflammation [647]. This is in line with the predictive value of TRAIL levels as a biomarker for heart failure in patients [648, 649]. Of note, TRAIL has also been reported to exert apoptosis-independent roles in cardiomyocyte growth and heart hypertrophy [650], as well as in angiogenesis and neovascularization upon experimental hindlimb ischemia [651]. Similar to neurological conditions, while TNF-R2 signaling appears to exert cardioprotective effects, the engagement of TNF-R1 drives cardiac hypertrophy, inflammation and cardiomyocyte loss [652,653,654,655,656,657,658,659]. The opposite outcome of TNF-R1 vs TNF-R2 signaling has been invoked to explain the clinical failure of TNF blocking agents in patients with chronic heart failure [660], despite encouraging preliminary findings [661, 662], as well as the cardiotoxic effects associated with the use of TNF blockers in patients with rheumatoid arthritis [663]. Confirming the involvement of extrinsic apoptosis in cardiac diseases, cardiomyocyte-specific deletion of Fadd in mice improved cardiomyocyte survival and heart function after ischemia/reperfusion [664]. Accordingly, haploinsufficiency of the gene encoding c-FLIP increased infarct area and aggravated cardiac dysfunction in mice experiencing myocardial infarction, while the cardiomyocyte-specific overexpression of c-FLIP attenuated pathology [665, 666]. Cardioprotection has also been observed in a mouse model of ischemia/reperfusion upon shRNA-mediated CASP8 depletion [667] or treatment with the CASP8 inhibitor Q-LETD-OPh [668]. Moreover, transplantation of Casp8−/− cells did not increase neovascularization in wild-type mice subjected to hindlimb ischemia [669], in line with a crucial role of CASP8 in the maintenance of endothelia in healthy conditions [596] (see Box 7). That said, combined pharmacological inhibition of apoptosis and necroptosis exerted greater cardioprotection than monotherapy in myocardial ischemia-reperfusion injury [670], suggesting the involvement of multiple RCD pathways in cardiovascular disorders.
FASL neutralization has been reported to improve cardiomyocyte survival and cardiac function in a model of cirrhotic cardiomyopathy [671]. Conversely, a cardioprotective effect of TRAIL and TNF was observed in mice developing cardiomyopathy upon the deletion of apolipoprotein E (ApoE) [672] or Des [673], respectively. Both FASL deficiency and administration of CASP8 inhibitors decrease tissue inflammation and aneurysm formation in mice subjected to CaCl2-induced abdominal aortic aneurysms [674]. A potential role of extrinsic apoptosis in gradual cardiomyocyte attrition during heart failure with reduced fraction has also been reported in a transgenic mouse model of inducible CASP8 overexpression [675]. Concerning TNF receptors, deletion of Tnfrsf1b resulted in increased cardiomyocyte death and hypertrophy induced by isoproterenol [676]. In contrast, deletion of Tnfrsf1a (but not Tnfrsf1b) was shown to be cardioprotective in murine models of vascular thrombosis [677], and heart failure based on angiotensin II administration [678]. Similar cardioprotection in this model has been reported after silencing of Tnfrsf1a [679]. In line with these findings, Cflar+/- mice (which lack one copy of the gene encoding c-FLIP) displayed increased sensitivity to cardiac injury upon angiotensin II administration [680].
FASL and TNF have also been reported to promote cardiac maladaptation and hypertrophy in models of pressure overload [681,682,683,684,685]. Consistent with this notion, TNF inhibition [686] or transgenic c-FLIP overexpression [687] limited experimental heart hypertrophy driven by hypertension. Moreover, treatment with etanercept reduced cardiac fibrosis in a diet-induced mouse model of obesity [688]. Conversely, both FAS and TNF receptor superfamily member 10b (TNFRSF10B, best known as TRAIL-R2 or mTRAIL-R) were reported to protect mice against atherosclerosis, at least in part by modulating TNF superfamily member 11 (TNFSF11, best known as RANKL) signaling [689,690,691,692,693], while the impact of TNF on experimental atherosclerosis remains a matter of debate [694,695,696,697]. Finally, pharmacological inhibition of TNF prevented cardiotoxicity induced by doxorubicin in mice [698,699,700]
Renal conditions
FASL, TNF and TRAIL have all been implicated in the development of acute kidney injury by driving the activation of both extrinsic apoptosis and inflammation. Loss-of-function mutations in Fasl, inhibition or depletion of FASL [701,702,703] as well as Fas [704] or Tnf [705] silencing, TNF neutralization [706, 707], or TRAIL blockade [708] exerted nephron-protective effects in mouse models of renal ischemia/reperfusion. Generation of chimeric mice reconstituted with spleen cells from gld/gld mice (lacking FASL) revealed a particular impact of FASL signaling in the hematopoietic compartment on ischemic acute kidney injury [702]. However, some functional overlap between DRs has also been reported. Indeed, while one study suggested that FASL neutralization was more effective than Tnfrsf1a deletion (leading to lack of TNF-R1) in preventing renal inflammation and cell death after acute kidney injury [701], another study reported that the neutralization of TNF but not FASL prevented tubular apoptosis and renal atrophy upon ischemia/reperfusion injury [706].
TRAIL blockade reportedly protects mice against renal damage after full-thickness scald burn, burn of all layers of the skin including epidermis and dermis [709], while TNF inhibition limited nephrotoxicity in mice treated with cisplatin [710], and acute tubulointerstitial nephritis in cancer patients administered with immune checkpoint inhibitors [711]. TNF neutralization also reduces tubulointerstitial fibrosis and renal injury in a mouse model of unilateral urethral obstruction [712, 713]. Contesting these findings, Tnf−/− mice showed increased fibrosis at later stages of ureteral obstruction [714]. This apparent discrepancy may reflect the distinct contribution of TNF-R1 and TNF-R2 signaling to different stages of renal fibrosis driven by urethral obstruction [715]. Conversely, experiments with lpr/lpr mice subjected to unilateral urethral ligation demonstrated a limited impact of FAS signaling to pathology [716]. The involvement of CASP8 in acute kidney injury is debated. While Casp8 and Casp3 deletion protected kidneys against damage induced by renal ischemia, increasing the survival of these mice [704, 717], such a nephroprotective effect was not observed after treatment with the broad-spectrum caspase inhibitor Z-VAD-FMK [718], potentially due to caspase inhibition promoting necroptosis after DR stimulation. In line with this notion, chemical inhibitors of receptor-interacting serine/threonine kinase 1 (RIPK1) as well as deletion of Ripk3 exerted robust nephroprotection in mouse models of ischemia/reperfusion [718, 719]. However, combined deletion of Casp8 and Ripk3 did not extend the beneficial effects of the genetic loss of Ripk3 and was associated with a more pronounced demise of tubular epithelial cells by intrinsic apoptosis [720].
DR activation has also been associated with chronic kidney disorders, but evidence involving CASP8-mediated apoptotic death is lacking. The conditional deletion of Tnf from macrophages [721], as well as the administration of TNF inhibitors [721,722,723,724], were reported to mediate beneficial effects in murine models of diabetic nephropathy. Conversely, the impact of TRAIL on this renal condition remains unclear [725,726,727], like that of TNF on polycystic kidney disease [728, 729]. As for glomerular inflammation, gld/gld mice (lacking FASL), as well as wild-type mice treated with TNF blockers, displayed reduced tissue damage during crescentic glomerulonephritis [730,731,732,733]. Indeed, balanced TNF-R1 and TNF-R2 signaling appeared to be critical for mice to resist experimentally induced glomerulonephritis [734,735,736,737,738,739]. This may explain apparently discrepant findings obtained with TNF-targeting measures.
Hepatic disorders
TNF-deficient mice, as well as rodents treated with TNF inhibitors, present with attenuated liver injury and apoptosis upon experimental ischemia/reperfusion, resulting in improved survival [740,741,742]. Of note, this beneficial effect cannot always be recapitulated in lpr/lpr and gld/gld mice, lacking FAS or FASL, respectively [742]. Similarly, FAS inhibition, FASL neutralization, as well as administration of low-dose TNF (as a pre-conditioning maneuver) have been shown to protect the liver against ischemia/reperfusion injury by reducing hepatic cell apoptosis and/or inflammation [743,744,745]. Protection of the liver from ischemia/reperfusion has also been observed in mice deficient for TRAIL [746], as well as upon the conditional knockdown of CASP8 or CASP3, the combined deletion of Casp8 and Ripk3, and the transgenic expression of a BID mutant that cannot be cleaved by CASP8 [283, 747, 748].
Lpr/lpr mice [749], Tnfsf10−/− mice (which lack TRAIL) [286], as well as animals exposed to TRAIL blockers [750], were protected against acetaminophen-induced liver damage, in line with the notion that FAS signaling and TRAIL receptor exacerbate acetaminophen hepatotoxicity [751]. Along similar lines, the hepatocyte-specific deletion of the gene encoding c-FLIP enhances liver injury and fibrosis induced by treatment with CCl4 or thioacetamide [752]. Moreover, a large body of evidence demonstrates that the abrogation of extrinsic apoptosis protects mice against fulminant hepatitis and hemorrhage in the liver induced by FASL and TNF. This has been achieved with strategies including (but not limited to) FADD blockade [753, 754], Casp8 [596, 755, 756] or Fadd [757] ablation, and Casp8 silencing [758]. Accordingly, hepatocyte-specific deletion of Cflar augments liver damage in mouse model of acute hepatic injury [759]. Consistent with the notion that engagement of the intrinsic apoptotic pathway is critical for DR induced cell killing in the liver, Bid−/− mice resist fatal hepatitis induced by FAS or TNF [278, 282, 760, 761], a protection that is enhanced by the concomitant loss of BIM or CASP8 [282]. Conditional deletion of the genes encoding BAX, BAK1 or PUMA, as well as overexpression of BCL2, can also protect hepatocytes from FAS-induced killing [762,763,764,765]. The impact of loss of BAD on TNF-induced hepatitis is controversial [766, 767]. Mice deficient for CASP3 or treated with CASP3 or CASP8 inhibitors have also been shown to be less sensitive to FAS-induced hepatocyte apoptosis [768, 769]. Of note, some degree of functional compensation between caspases and alternative mechanisms of caspase activation have emerged from studies in hepatocytes responding to FAS agonists [770]. Finally, FAS and TNF-R1 signaling, as well as FADD activation, are involved in liver regeneration following partial hepatectomy [771,772,773,774,775]. In this context, the liver-specific deletion of Casp8 resulted in dysregulated hepatocyte proliferation upon partial hepatectomy coupled to the initiation of an inflammatory response [776]. It has been suggested that CASP8 modulates liver regeneration by balancing NF-κB activation and necroptosis rather than by inducing apoptosis [777].
Gld/gld mice (lacking FASL) chronically fed with ethanol display reduced liver injury, steatosis and inflammation as compared to wild-type mice, but exhibit signs of incipient fibrosis [778]. Some degree of protection against alcohol-induced liver damage has also been documented in mice deficient for the apoptosis-inducing TRAIL receptor mTRAIL-R [779] or TNF-R1 (but not TNF-R2) [780], as well as in mice receiving a TRAIL-neutralizing antibody [781]. Accordingly, the hepatocyte-specific ablation of Casp8 limited hepatic steatosis in murine models of ethanol administration, although it failed to prevent apoptotic RCD [782]. Conversely, apoptosis driven in hepatocytes by chronic ethanol exposure could be abolished by systemic inhibition of CASP3 with Ac-DEVD-FMK [783].
The liver-restricted overexpression of FAS induces hepatic steatosis and insulin resistance in mice subjected to a high-fat diet (HFD) [784]. In the same experimental setting, hepatoprotection was observed with the hepatocyte-specific ablation of Fas or germline deletion of Bid [784]. Moreover, Tnf deletion [785, 786], whole-body deletion of Tnfrsf1a (encoding TNF-R1) alone or in combination with the gene encoding TNF-R2 [787, 788] as well as inhibition of TNF [789,790,791] or TNF-R1 [792] significantly reduced hepatic steatosis, fibrosis, damage, and metabolic alterations in several diet-induced or genetic models of non-alcoholic fatty liver disease (NAFLD). In apparent contrast with these findings, the hepatocyte-specific deletion of Tnfrsf1a failed to protect mice from diet-driven NASH [793]. Moreover, Tnfrsf1a deletion accelerated the progression of steatosis to steatohepatitis in mice on a HFD [794]. Taken together, these findings underscore the pleiotropic and context-dependent effects of TNF/TNF-R signaling in NAFLD. The impact of TRAIL on NAFLD is also debated. Indeed, contrasting evidence from experiments with mice deficient for TRAIL or treated with recombinant TRAIL suggests either a detrimental or a beneficial role to TRAIL in NAFLD induced by HFD [795,796,797].
The absence of mTRAIL-R promoted hepatic inflammation and fibrosis in a genetic mouse model of cholestasis [798]. Similarly, lpr/lpr mice lacking FAS [799,800,801] as well as TNF-deficient [802, 803] and TRAIL-deficient [804, 805] mice displayed reduced hepatocyte apoptosis and fibrogenesis after experimental cholestasis induced by bile duct ligation. In line with these results, expression of a phosphorylated FADD mimicking mutant attenuated HFD-induced hepatomegaly and steatosis [806]. Experiments based on the hepatocyte-specific deletion of Cflar (encoding c-FLIP) or transgenic overexpression of c-FLIP revealed a role for this modulator of CASP8 activation as a suppressor of hepatic steatosis and inflammation induced by HFD [807]. Moreover, the hepatocyte-specific deletion of Cflar in mice resulted in enhanced cholestatic liver injury and inflammatory responses upon bile duct ligation [808]. Similarly, the hepatocyte-specific deletion of Casp8 protected mice against liver injury in models of cholestatic hepatitis caused by the administration of 3,5-diethoxycarbonyl-1,4-dihydrocollidine [809], as well as in models of steatosis caused by the feeding of a methionine- and choline-deficient diet [810]. A comparable hepatoprotection against obstructive cholestasis has been documented in mice with hepatocyte-specific Casp8 deletion [811]. Furthermore, liver parenchymal cell-specific ablation of the gene encoding FADD prevented RIPK1-dependent but not TNF-R1-, FAS-, and TRAIL-R-dependent hepatocyte apoptosis, chronic liver inflammation and hepato-carcinogenesis in mice with liver-specific deficiency in inhibitor of kappaB kinase gamma (IKBKG, best known as NEMO or IKKgamma) [812, 813]. Finally, decreased BID cleavage has been associated with attenuated liver injury in mouse models of chronic cholestasis [814].
Hematologic malignancies and solid cancers
Human patients with autoimmune lymphoproliferative syndrome (ALPS) caused by defects in FAS are known to show abnormally increased predisposition to lymphoma development [815]. Accordingly, FAS-deficient lpr/lpr mice develop a plasmacytoma-like disease in advanced age [816]. TRAIL also seems to exert a tumor suppressive function in lymphomagenesis. The ablation of the gene encoding mTRAIL-R accelerated the development of lymphoma in Eμ-Myc transgenic mice [817]. Moreover, deficiency in TRAIL (but not in mTRAIL-R) promoted the development of lymphoma and other tumors in mice with haploinsufficiency for Trp53 [818, 819]. Interestingly, mice engineered to express exclusively either membrane-bound or secreted FASL showed an increased incidence of spontaneous tumor formation when expressing only soluble FASL, which is unable to induce FAS-mediated apoptosis but may exert pro-inflammatory effects [820].
The role of FAS and TRAIL-R in the development of colorectal cancer is controversial. For instance, the loss of FAS was reported to enhance Apc mutation-induced but not inflammation-induced intestinal tumorigenesis [821,822,823]. Along similar lines, while the ablation of Tnfrsf10b (leading to lack of mTRAIL-R) in mice did not impact tumorigenesis induced by Apc mutations [819], the administration of TRAIL suppressed tumorigenesis in a mouse model of colitis-associated colon cancer [824]. Despite some contention in this respect [825,826,827,828], TNF seems to contribute to the development of colorectal cancer, although whether such effects depend on the apoptotic function of TNF needs to be formally established. The administration of TNF blockers [829,830,831,832,833] or ablation of Tnf [834] or Tnfrsf1a [834, 835] limits colorectal oncogenesis, as shown in animal models of colorectal cancer induced by colitis, chemicals, or mutations in Apc. Finally, loss of the dependence receptor DCC netrin 1 receptor (Dcc) accelerates cancer progression in a mouse model of Apc mutation-driven colorectal oncogenesis [836]. A tumor suppressor role in colorectal cancer has also been described for the dependence neurotrophic tyrosine kinase, receptor, type 3 (NTRK3, best known as TRKC) [837]. Of note, the association between gain of dependence receptors ligands (e.g., NTN1) with tumor progression [838], may make their targeting a promising anti-cancer approach [839].
With regard to other tumor types, both TNF-R1 and FAS display a pro-oncogenic role in hepatic and ovarian oncogenesis. Specifically, conditional deletion of Fas in hepatocytes delays carcinogen-driven hepatocarcinogenesis, while Fas ablation suppresses the development of ovarian tumors in phosphatase and tensin homolog (PTEN)-deficient mice bearing mutant Kirsten rat sarcoma viral oncogene (Kras) [840]. Likewise, TNF neutralization limits the onset of hepatic cancer driven by experimentally induced cholestatic hepatitis [841]. Consistent with these findings, Casp8−/− mice are protected against the development of inflammation-driven liver cancer [755]. Hyperactivation of CASP8 in the context of RIPK1 and TNF receptor-associated factor 2 (TRAF2) deficiency has indeed been implicated in the development of hepatocellular carcinoma [842], although such effects may be independent of apoptosis induction [843, 844]. In contrast, recent studies show a tumor-suppressive function of CASP8 in the liver and certain other tissues [845,846,847,848]. In particular, there is evidence of a role of CAPS8 in early tumorigenesis (but not tumor progression) exerted by modulating the DNA damage response [845] or the level of chromosomal instability (CIN) [846].
Consistent with a pro-tumorigenic effect of TNF, the ablation of Tnf or Tnfrsf1a or the blockade of TNF in mice conferred protection against carcinogen-induced skin oncogenesis [849,850,851,852,853,854]. In contrast, the impact of genetic and pharmacological inhibition of TNF in UVB-induced skin cancer is debated [855, 856]. Of note, the comparison between TNF-R1- vs. TNF-R2-deficient mice revealed a primary role of TNF-R1 in chemically induced skin oncogenesis [851]. Furthermore, TNF-R1 deficiency suppresses the development of skin cancer induced by NF-κB inhibition [857]. A similar role for TNF-R1 in supporting tumorigenesis was described in murine models of N-methyl-N-nitrosourea(NMU)/testosterone-induced prostate cancer [858] and methylcholanthrene (MCA)-induced fibrosarcoma [859]. As opposed to TNF-R1, TNF-R2 shows tumor-suppressive functions in mouse models of tumorigenesis, such as the development of fibrosarcoma triggered by MCA [859], and mammary oncogenesis induced by transgenic expression of wingless-type MMTV integration site family, member 1 (Wnt1) [860]. Moreover, the absence of TNF impairs tumor growth in HER2-driven mammary tumorigenesis in mice [861], and TNF neutralization suppresses chemically induced oral [862] and pulmonary [863] tumorigenesis. Conversely, TNF overexpression in the airway epithelium enhanced mutant Kras-driven lung oncogenesis [864].
Pre-clinical evidence points to some tumor type-specificity for the role of TRAIL and its receptor(s) in tumorigenesis. Transgenic expression of TRAIL in the skin delays chemically induced carcinogenesis [865]. This effect can be recapitulated in mice lacking TRADD [866] but, curiously, not in mTRAIL-R-deficient mice [867], with the latter actually showing enhanced lymph node involvement. In support of an anti-tumor function for the TRAIL/TRAIL-R system, TRAIL-deficient mice as well as mice treated with TRAIL blockers displayed increased susceptibility to MCA-induced fibrosarcoma [868, 869]. In a recent study, administration of recombinant TRAIL coupled to inhibition of cyclin-dependent kinase 9 (CDK9) was effective in a wide range of cancers [870]. Yet in contrast to this and in support of a tumor-supportive role of endogenous TRAIL, deficiency in mTRAIL-R limits tumor growth and improves survival in mouse models of mutant Kras-driven lung and pancreatic tumorigenesis [871]. Moreover, malignant cell-specific ablation of genes encoding mTRAIL-R or FADD reduced lung cancer growth and tumor-promoting inflammation [872], while systemic ablation of Tnfsf10 (leading to lack of TRAIL) had no impact on HER2-driven mammary oncogenesis [818]. Interestingly, KRAS mutations have been shown to promote the switch of FAS and TRAIL receptors from a predominantly death-inducing into a metastasis promoting function [873]. Since TRAIL as well as FASL can trigger either apoptosis, necroptosis, inflammation or pro-invasive signaling, cancer-specific preferences for one or the other of these outputs are likely accounts for the apparently discrepant effects observed in various cancer models.
Autoimmune and inflammatory diseases
The interpretation of results on the impact of extrinsic apoptosis in the etiology of autoimmune and inflammatory disease should consider the fact that DR engagement can also result in the initiation of an inflammatory response not related to RCD (see Box 6 and Box 7), meaning that DR deregulation may lead to inflammatory diseases independently of the induction of extrinsic apoptosis. The notion that defects in DR signaling can cause autoimmune disease is supported by the observation that lpr/lpr as well as gld/gld mutant mice, deficient for FAS or FASL, respectively, as well as humans with defects in FAS develop an SLE-like autoimmune disease accompanied by lymphadenopathy, splenomegaly and hepatomegaly [874, 875]. A critical role for loss of CASP8-mediated apoptosis in this disease was demonstrated by the observation that a similar condition is seen in mice lacking Casp8 and also Ripk3 or Mlkl (to prevent necroptosis) [51, 52, 876]. However, the roles of DRs in autoimmune disease are complex. TRAIL/TRAIL-R signaling was reported to protect mice and rats against autoimmune encephalomyelitis [877,878,879,880,881,882], autoimmune arthritis [883,884,885,886,887] and type I diabetes [690, 883, 888,889,890,891]. Perhaps surprisingly, the presence of FAS and TNF-R1 is associated with the development of certain autoimmune conditions. Indeed, both lpr/lpr lacking FAS and gld/gld mice lacking FASL, as well as TNF-R1-deficient mice, appear to be protected against experimental encephalomyelitis [892,893,894,895]. Similar results were obtained in mice with Tnf deletion in monocytes and macrophages, but not in mice lacking TNF in microglial cells [896]. Protection against experimentally induced autoimmune conditions were also found in mice subjected to neutralization of TNF or TNF-R1 inhibition [897,898,899,900,901,902,903,904]. FAS-independent mechanisms also appear to support the pathogenesis of experimental autoimmune encephalomyelitis [892, 905], with some studies pointing to a protective role for FAS-induced RCD amongst lymphocytes in this disease model [906]. Moreover, FAS engagement was reported to differentially contribute to the initiation vs. the recovery from autoimmune encephalomyelitis [907, 908]. In particular, FASL expression in astrocytes appears to promote recovery from experimental autoimmune encephalomyelitis, as shown by persisting demyelination and paralysis of mice with an astrocyte restricted deletion of Fasl [907]. Finally, at least in some studies, Tnf deletion or TNF neutralization failed to attenuate the severity of autoimmune encephalomyelitis once the disease was established [909, 910].
Mice with defects in FASL or TNF signaling are protected against arthritis induced by immunization with xenogeneic type II collagen in complete Freund’s adjuvant [911,912,913,914]. Similar protection was observed in mice transplanted with mesenchymal stem cells engineered to express TNF inhibitors [915]. In keeping with this evidence, the myeloid cell specific deletion of Fas or the administration of antibodies that target both TNF and chemokine (C-X-C motif) ligand 10 (CXCL10) resulted in accelerated disease resolution in a model of rheumatoid arthritis induced by K/BxN serum transfer [916, 917]. Genetic loss of Fas or pharmacological inhibition of FAS conferred protection against autoimmune diabetes in specific animal models, including NOD mice [918,919,920,921,922,923]. However, whether the impact of FAS on the pathogenesis of autoimmune diabetes depends on its role in the death of pancreatic β-cell [918] or its role in inflammation (e.g., in the context of insulitis) remains a matter of debate [921]. Other studies found no role for FAS in diabetes [924,925,926]. TNF neutralization is effective only in a limited sub-group of patients with inflammatory bowel disease [927, 928]. This is in line with the finding that deletion of the gene encoding TNF-R1 exacerbated colitis in IL10-deficient mice [929]. A similar protection was ascribed to TRAIL/TRAIL-R signaling in a dextran sodium sulfate-induced model of colitis [930, 931]. Finally, it has been suggested that FASL and TNF signaling contribute to the pathogenesis of acute pancreatitis [932, 933]. A similar detrimental role has been proposed for TNF in autoimmune neuritis [934,935,936], although there is also some contention [937], as well as in spondylarthritis [938] and psoriasis [939]. Conversely, mTRAIL-R appears to mediate beneficial effects in autoimmune thyroiditis [940,941,942,943,944] At least in part, these findings reflect the pleiotropic effects of whole-body inhibition of DRs signaling, which concomitantly impacts both the target (i.e., parenchymal) and the perpetrator (i.e., immune cells) of damage.
Some experimental evidence links CASP8 activation to autoimmune and inflammatory disorders. In a recent study using a chemically-induced model of intestinal inflammation, the selective absence of CASP8 in intestinal epithelial cells decreased their survival, also resulting in gut barrier dysfunction and chronic inflammation [945]. Of note, in this setting, inflammation can occur via a mechanism independent of the induction of necroptosis (which is inhibited by CASP8) and involving the activation of RIPK1 and RNA sensor RIG-I (RIGI; best known as RIG-I) [946, 947]. Along similar lines, chronic proliferative dermatitis in mice deficient for components of the linear ubiquitin chain assembly complex (LUBAC) has been associated with an increased keratinocyte apoptosis mediated by the engagement of TNF-R1 and the activation of the RIPK1- and/or FADD-CASP8 cascade [948,949,950,951,952]. Importantly, in this mouse model of an inflammatory disease the relevant contributions of cell death versus inflammatory signalling from TNF-R1 were genetically dissected demonstrating that excess apoptosis/necroptosis drove different elements of the inflammatory response depending on the affected tissue. In a mouse model of autoimmune encephalomyelitis, the oligodendrocyte-specific deletion of Fadd reduced demyelination and this was accompanied by limited immune cell infiltration in the spinal cord [953]. Likewise, experimental autoimmune encephalomyelitis could be prevented by transgenic expression of FADD-DN (dominant negative form of FADD) in T cells [954], but it must be noted that this reflects the death of activated T cells [955]. Therefore, this protective effect is due to the removal of T cells that would cause tissue destruction. Activation of CASP8 was identified in the microglia of patients with multiple sclerosis [956]. Moreover, transgenic expression of FADD-DN or Casp8 ablation in pancreatic β cells protects mice from autoimmune diabetes [957]. Finally, BID appears to be dispensable for the development of diabetes in NOD mice [958].
There are also contrasting observations on the impact of DR-induced apoptosis on the development and resolution of autoimmune rheumatoid arthritis. The absence of c-FLIP (due to Cflar deletion) increased disease severity but limited disease resolution in mice experiencing arthritis upon intraperitoneal injection of serum from K/BxN mice [959]. In the same model, deletion of Casp8 in all myeloid cells enhanced disease resolution, while deletion of Casp8 selectively in DCs accelerated disease onset [960]. Further experiments are required to unveil the reasons for such cell type specificity in the role of CASP8 in this and (perhaps) other autoimmune disorders.
Infectious diseases
Extrinsic apoptosis is reported to act as an anti-infective mechanism. FAS-deficient lpr/lpr, FASL-deficient gld/gld and Bid−/− mice exhibit delayed clearance of Citrobacter rodentium and increased intestinal pathology [961]. Confirming the importance of DR-induced apoptosis, this pathogen was shown to inhibit extrinsic apoptosis of infected enterocytes by expressing specific virulence proteins, such as the N-acetylglucosamine transferase NleB1, which prevents FADD-mediated recruitment and activation of CASP8 [962]. Along similar lines, Fas−/− mice have shorter lifespan than wild-type mice after challenge with L. monocytogenes, succumbing to neurolisteriosis. This was proposed to be promoted by an impaired loss of monocytes due to upregulated expression of c-FLIP by the bacterial protein InlB [963]. In support of this result, conditional deletion of Cflar in myeloid cells improved L. monocytogenes clearance and host survival [964]. FAS signaling also conferred protection from infection with (i) human herpes simplex virus 2 (HSV-2), as demonstrated by a decrease in the loss of monocyte and immune cell recruitment at the infection site in Fas−/− and Fasl−/− mice [965], and (ii) C. rodentium or lymphocytic choriomeningitis virus, as demonstrated by an increased neutrophil fraction in mice with conditional deletion of Fas in the myeloid compartment [966].
Supporting an anti-infection role of CASP8, mice lacking RIPK1 kinase activity fail to control systemic Yersinia infection, rapidly dying because of excess apoptosis driven by a kinase-independent function of RIPK1 [967, 968]. In line with this finding, Ripk3−/−Casp8−/− (but not Ripk3−/−) mice die from Toxoplasma gondii infection due to acute toxoplasmosis [969]. Moreover, the hepatocyte-specific deficiency for CASP8 facilitates mouse liver infection by L. monocytogenes, resulting in inflammation and development of necrotic lesions [776]. These results also suggest an interconnection of multiple RCD pathways in controlling infection. Indeed, the deletion of Z-DNA binding protein 1 (Zbp1), an essential cytoplasmic sensor of Influenza A virus (IAV) Z-RNA required for the activation of MLKL-dependent necroptosis, RIPK1/FADD-dependent apoptosis and NLR family, pyrin domain containing 3 (NLRP3) inflammasome-dependent pyroptosis, as well as co-deletion of the genes encoding MLKL and FADD, causes a defect in the control of IAV infection and lethal respiratory failure. These findings support an essential role of apoptosis, necroptosis and pyroptosis in IAV clearance [970,971,972,973,974]. Similarly, combined activation of apoptosis and other RCD pathways contribute to the response of mice to Burkholderia thailandensis infection [975]. Finally, pharmacological or tissue specific genetic deletion of baculoviral IAP repeat-containing 3 (Birc3, encoding cIAP1) and baculoviral IAP repeat-containing 2 (Birc2, encoding cIAP2) results in better control of hepatitis B virus and liver stage malaria parasites due to increased TNF induced death of infected cells [976,977,978].
Experimental evidence also suggests a detrimental role of extrinsic apoptosis during some infections. Mice deficient for both TNF-R1 and TNF-R2 display decreased sensitivity to LPS, suggesting a critical role for TNF in tissue injury during gram-negative bacterial infection [979]. Along similar lines, TNF-R1-deficient mice are more resistant than wild-type mice to the cytopathic effects of TNF during Sindbis virus infection, as evidenced by delayed paralysis and reduced mortality [980]. Moreover, ablation of Ripk1 protected mice from acute liver injury after infection with L. monocytogenes [981], while knockout of Fas or Fasl reduced the effect of toxin A-induced enteritis in mice infected with Clostridium difficile, which has been attributed to a reduction in enterocyte loss [982]. Additionally, the infectious spleen and kidney necrosis virus (ISKNV) induced tissue damage in zebrafish by activation of DR-induced apoptosis by a viral protein encoding a TRADD interactor [983]. Of note, in this study, the absence of CASP8 protected zebrafish from ISKNV infection. Finally, Ripk3−/−Casp8−/− mice exhibit high levels of protection from LPS-induced septic shock [984] or a lethal cytokine shock and tissue damage driven by TNF and interferon gamma (IFNG), mirroring that of SARS-CoV-2 [985]. This suggests that the several types of RCD can mediate infection-associated pathogenesis, as demonstrated for infection with Salmonella [50].
Other diseases
TNF is reported to impair myogenesis in a mouse model of skeletal muscle regeneration after hindlimb immobilization (hindlimb suspension) [986]. Moreover, silencing of TRAIL improved muscle regeneration in mice with acute skeletal muscle injury due to local injection of BaCl2 [987]. An inhibitory role in myogenesis has also been ascribed to FADD, at least in response to freezing-induced muscle injury [988]. In apparent contrast with this result, combined deletion of the genes encoding TNF-R1 and TNF-R2 limited skeletal muscle regeneration after cardiotoxin-induced injury [989, 990], suggesting the relevance of a balance between TNF-R1 and TNF-R2 signaling in this model. TRAIL neutralization increased muscular strength in a mouse model of Duchenne muscular dystrophy [991], while other studies associated TRAIL and FASL signaling to myositis [992, 993].
Activation of DRs has also been implicated in the pathogenesis of acute lung injury. Fas silencing as well as TNF neutralization protected mice from lung injury induced by ischemia-reperfusion [994, 995]. Similarly, deletion of Tnfrsf1a (encoding TNF-R1) or pharmacological inhibition of TNF-R1 or CASP8 attenuated pulmonary edema formation and improved alveolar epithelial function in a murine model of acute lung injury induced by acid inhalation [996, 997]. A similar protective effect was provided by pharmacological inhibition or genetic deletion of FASL or TNF in a LPS-induced mouse model of acute lung injury [998,999,1000,1001,1002,1003,1004]. However, in one study FAS signaling was shown to contribute to the resolution of acute lung injury by depleting macrophages [1005]. Using distinct mouse models of acute lung damage following sepsis, it has been shown that abrogation of FAS and TNF-R1 signaling decreases pulmonary apoptosis and ameliorates pathology, with a survival benefit in some settings [1006,1007,1008,1009,1010,1011,1012]. Hyperoxia-induced lung injury and bleomycin-induced pulmonary fibrosis, a model for cancer therapy-induced lung injury, are also impacted by DR signaling. Thus, FAS and TNF deficiency exacerbated hyperoxia-induced lung injury and/or inflammation in newborn mice [1013, 1014]. In contrast, TNF inhibition conferred protection against hyperoxia-induced lung damage in a murine model [1015,1016,1017]. Moreover, the absence of TNF-R1 (but not TNF-R2) improved survival in mice subjected to excessive oxygen supply, and this was not linked to decreasing inflammation [1018]. In support of these results, specific ablation of Fas in murine fibroblasts or T cells exacerbates pulmonary fibrosis induced by bleomycin [1019, 1020]. However, the level of bleomycin-induced pulmonary fibrosis is reduced in FAS-deficient lpr/lpr or FASL-deficient gld/gld mice [1021], but remains unchanged in mice treated with FAS-neutralizing agents [1022]. Likewise, contrasting findings support or refute a role for TNF [1023,1024,1025] and TRAIL [1026, 1027] in the onset and resolution of pulmonary fibrosis after administration of bleomycin. TNF neutralization has been reported to attenuate and enhance interstitial pulmonary fibrosis induced by nitrogen mustard [1028] or rituximab [1029]. Finally, FASL, TNF and/or TRAIL have been implicated in infectious or non-infectious lung disorders, including (but not limited to) infection with respiratory syncytial virus (RSV) [1030,1031,1032,1033,1034,1035,1036], adenovirus type 1 respiratory disease [1037, 1038], allergic reaction and asthma [1039,1040,1041,1042,1043,1044,1045,1046,1047,1048,1049,1050] and idiopathic pneumonia syndrome [1051], as well as to chronic lung diseases (e.g., chronic obstructive pulmonary disease) [864, 1052, 1053].
The studies discussed above illustrate that DR-induced apoptosis is at the heart of many disorders either promoting recovery or exacerbating disease. The active involvement in disease severity and progression makes this pathway a potentially tractable target for therapeutic interventions in a wide range of diseases, typically those with an inflammatory component. However, this effect may be linked to the role of DR signaling in other RCD pathways and in inflammation. Moreover, there is little consensus on the roles of FASL, TNF and/or TRAIL in these pathologies, highlighting a high complexity of the system that calls for further investigation.
Concluding remarks
Abundant preclinical evidence demonstrates that the intrinsic and the extrinsic pathways of apoptosis not only contribute to adult tissue homeostasis and, in the case of the intrinsic pathway, to embryonic development – the implication of CASP8 in development is mainly linked to its role as necroptosis inhibitor (see Box 6 and Box 7) - but also contribute to the pathogenesis of multiple diseases, including various cardiovascular, hepatic, neurological and renal disorders as well as multiple infectious, autoimmune, inflammatory and oncological conditions. However, despite great potential as targets for therapeutic interventions and a considerable research effort dedicated to developing effective approaches, the success of intrinsic or extrinsic apoptosis-targeting agents in clinical settings is so far limited to BH3 mimetic drugs, SMAC mimetics, caspase inhibitors as well as activators or inhibitors of DR signaling, with only one compound, the BCL2 inhibitor venetoclax being approved for the treatment of patients with CLL or AML.
Rather than mitigating the enthusiasm about the clinical potential of modulators of apoptosis, this challenge suggests the need for a substantial change in the experimental design and re-interpretation of results, at different levels (Fig. 1). One major issue is that studies evaluating the impact of apoptotic cell death on disease have not always addressed the connections between the core components of the intrinsic and extrinsic apoptotic machinery or their potential interaction and functional overlap with other RCD pathways. Also, the potential activation of alternative RCD modalities as a mechanism to compensate for the inhibition of apoptotic RCD has not always been explored as an approach to achieve superior outcomes. The importance of independent replication of findings that suggest success from targeting a pathway in the treatment of a disease cannot be emphasized enough. Only then can the costly process of clinical translation be approached with confidence and with an increased chance of success. For example, the findings that overexpression of BCL2 or its pro-survival relatives can promote tumorigenesis and can render malignant cells resistant to diverse anti-cancer therapeutics had been reproduced hundreds of times before the initiation of BH3 mimetic development. This is not yet the case for many of the other studies discussed herein, as best demonstrated by the fact that certain experiments have provided diametrically opposing results in different laboratories. These questions must be resolved before considering novel drug development programs around apoptotic RCD.
Moreover, some regulators of apoptosis and signaling cascades have been reported to exert a variety of functions beyond cell death control, including (but not limited to) inflammation (e.g., multiple caspases and IAPs), cell differentiation (e.g., pro-and anti-apoptotic BCL2 proteins), cell proliferation and survival (e.g., DR engagement). The relevance of these functions is often dependent on cell/tissue type (as it is related to variable expression levels and activation status of other regulators of RCD) and the intensity and duration of the initiating stimulus (as they can direct to a distinct biological outcome, as exemplified by DR ligation). Of note, some of these cell death unrelated functions of bona fide cell death regulators are highly controversial and much more work must be done to verify or discard them. On the one hand, this pleiotropy may result in a variable (even including an antagonistic protective vs. promoting) impact of apoptosis on distinct human diseases, which may also explain the considerable degree of context-dependency observed during its experimental modulation. On the other hand, the pathogenic effect of core components of the apoptotic machinery is often mediated by apoptosis-unrelated functions including inflammation, which may point to unexplored targets for the development of new therapeutic approaches.
In our opinion, investigating the molecular cascade of apoptotic cell death in the context of the functional interconnection between apoptotic and non-apoptotic RCD pathways, for instance by interrupting some of the molecular connections between different RCD signaling cascades, may instigate new advances, ultimately leading to the development of novel, clinically-viable apoptosis modulators for use in multiple disease settings.
References
Galluzzi L, Vitale I, Aaronson SA, Abrams JM, Adam D, Agostinis P, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018;25:486–541.
Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021;22:266–82.
Del Re DP, Amgalan D, Linkermann A, Liu Q, Kitsis RN. Fundamental mechanisms of regulated cell death and implications for heart disease. Physiol Rev. 2019;99:1765–817.
Bock FJ, Tait SWG. Mitochondria as multifaceted regulators of cell death. Nat Rev Mol Cell Biol. 2020;21:85–100.
Broz P, Pelegrín P, Shao F. The gasdermins, a protein family executing cell death and inflammation. Nat Rev Immunol. 2020;20:143–57.
Weinlich R, Oberst A, Beere HM, Green DR. Necroptosis in development, inflammation and disease. Nat Rev Mol Cell Biol. 2017;18:127–36.
Galluzzi L, Buqué A, Kepp O, Zitvogel L, Kroemer G. Immunogenic cell death in cancer and infectious disease. Nat Rev Immunol. 2017;17:97–111.
Tang D, Kang R, Berghe TV, Vandenabeele P, Kroemer G. The molecular machinery of regulated cell death. Cell Res. 2019;29:347–64.
Gudipaty SA, Conner CM, Rosenblatt J, Montell DJ. Unconventional ways to live and die: cell death and survival in development, homeostasis, and disease. Annu Rev Cell Dev Biol. 2018;34:311–32.
Bedoui S, Herold MJ, Strasser A. Emerging connectivity of programmed cell death pathways and its physiological implications. Nat Rev Mol Cell Biol. 2020;21:678–95.
Galluzzi L, Vitale I, Abrams JM, Alnemri ES, Baehrecke EH, Blagosklonny MV, et al. Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ. 2012;19:107–20.
Kesavardhana S, Malireddi RKS, Kanneganti TD. Caspases in cell death, inflammation, and pyroptosis. Annu Rev Immunol. 2020;38:567–95.
Kumar S, Dorstyn L, Lim Y. The role of caspases as executioners of apoptosis. Biochem Soc Trans. 2022;50:33–45.
Galluzzi L, Bravo-San Pedro JM, Vitale I, Aaronson SA, Abrams JM, Adam D, et al. Essential versus accessory aspects of cell death: recommendations of the NCCD 2015. Cell Death Differ. 2015;22:58–73.
Marsden VS, O’Connor L, O’Reilly LA, Silke J, Metcalf D, Ekert PG, et al. Apoptosis initiated by Bcl-2-regulated caspase activation independently of the cytochrome c/Apaf-1/caspase-9 apoptosome. Nature. 2002;419:634–7.
Cheng EH, Wei MC, Weiler S, Flavell RA, Mak TW, Lindsten T, et al. BCL-2, BCL-X(L) sequester BH3 domain-only molecules preventing BAX- and BAK-mediated mitochondrial apoptosis. Mol Cell. 2001;8:705–11.
Pandian N, Kanneganti TD. PANoptosis: a unique innate immune inflammatory cell death modality. J Immunol. 2022;209:1625–33.
Bonora M, Giorgi C, Pinton P. Molecular mechanisms and consequences of mitochondrial permeability transition. Nat Rev Mol Cell Biol. 2022;23:266–85.
Chen X, Zeh HJ, Kang R, Kroemer G, Tang D. Cell death in pancreatic cancer: from pathogenesis to therapy. Nat Rev Gastroenterol Hepatol. 2021;18:804–23.
Song X, Zhu S, Xie Y, Liu J, Sun L, Zeng D, et al. JTC801 Induces pH-dependent death specifically in cancer cells and slows growth of tumors in mice. Gastroenterology. 2018;154:1480–93.
Tsvetkov P, Coy S, Petrova B, Dreishpoon M, Verma A, Abdusamad M, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. 2022;375:1254–61.
Malireddi RKS, Kesavardhana S, Kanneganti TD. ZBP1 and TAK1: master regulators of NLRP3 inflammasome/pyroptosis, apoptosis, and necroptosis (PAN-optosis). Front Cell Infect Microbiol. 2019;9:406.
Green DR. The coming decade of cell death research: five riddles. Cell. 2019;177:1094–107.
Singh R, Letai A, Sarosiek K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol. 2019;20:175–93.
Ke FFS, Brinkmann K, Voss AK, Strasser A. Some mice lacking intrinsic, as well as death receptor induced apoptosis and necroptosis, can survive to adulthood. Cell Death Dis. 2022;13:317.
Ke FFS, Vanyai HK, Cowan AD, Delbridge ARD, Whitehead L, Grabow S, et al. Embryogenesis and adult life in the absence of intrinsic apoptosis effectors BAX, BAK, and BOK. Cell. 2018;173:1217–1230.e1217.
Spetz J, Galluzzi L. Preface: life through death-key role of cellular suicide for colonial and organismal homeostasis. Int Rev Cell Mol Biol. 2020;352:xi–xv.
Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020;17:395–417.
Anderton H, Wicks IP, Silke J. Cell death in chronic inflammation: breaking the cycle to treat rheumatic disease. Nat Rev Rheumatol. 2020;16:496–513.
Li K, van Delft MF, Dewson G. Too much death can kill you: inhibiting intrinsic apoptosis to treat disease. EMBO J. 2021;40:e107341.
Jain N, Keating M, Thompson P, Ferrajoli A, Burger J, Borthakur G, et al. Ibrutinib and Venetoclax for First-Line Treatment of CLL. N Engl J Med. 2019;380:2095–103.
Souers AJ, Leverson JD, Boghaert ER, Ackler SL, Catron ND, Chen J, et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat Med. 2013;19:202–8.
Diepstraten ST, Anderson MA, Czabotar PE, Lessene G, Strasser A, Kelly GL. The manipulation of apoptosis for cancer therapy using BH3-mimetic drugs. Nat Rev Cancer. 2022;22:45–64.
Merino D, Kelly GL, Lessene G, Wei AH, Roberts AW, Strasser A. BH3-Mimetic drugs: blazing the trail for new cancer medicines. Cancer Cell. 2018;34:879–91.
Fischer K, Al-Sawaf O, Bahlo J, Fink AM, Tandon M, Dixon M, et al. Venetoclax and obinutuzumab in patients with CLL and coexisting conditions. N Engl J Med. 2019;380:2225–36.
Roberts AW, Davids MS, Pagel JM, Kahl BS, Puvvada SD, Gerecitano JF, et al. Targeting BCL2 with venetoclax in relapsed chronic lymphocytic leukemia. N Engl J Med. 2016;374:311–22.
Seymour JF, Kipps TJ, Eichhorst B, Hillmen P, D’Rozario J, Assouline S, et al. Venetoclax-rituximab in relapsed or refractory chronic lymphocytic leukemia. N Engl J Med. 2018;378:1107–20.
DiNardo CD, Jonas BA, Pullarkat V, Thirman MJ, Garcia JS, Wei AH, et al. Azacitidine and Venetoclax in Previously Untreated Acute Myeloid Leukemia. N Engl J Med. 2020;383:617–29.
Frenette CT, Morelli G, Shiffman ML, Frederick RT, Rubin RA, Fallon MB, et al. Emricasan improves liver function in patients with cirrhosis and high model for end-stage liver disease scores compared with placebo. Clin Gastroenterol Hepatol. 2019;17:774–783.e774.
Garcia-Tsao G, Bosch J, Kayali Z, Harrison SA, Abdelmalek MF, Lawitz E, et al. Randomized placebo-controlled trial of emricasan for non-alcoholic steatohepatitis-related cirrhosis with severe portal hypertension. J Hepatol. 2020;72:885–95.
Harrison SA, Goodman Z, Jabbar A, Vemulapalli R, Younes ZH, Freilich B, et al. A randomized, placebo-controlled trial of emricasan in patients with NASH and F1-F3 fibrosis. J Hepatol. 2020;72:816–27.
Galluzzi L, Aaronson SA, Abrams J, Alnemri ES, Andrews DW, Baehrecke EH, et al. Guidelines for the use and interpretation of assays for monitoring cell death in higher eukaryotes. Cell Death Differ. 2009;16:1093–107.
Boada-Romero E, Martinez J, Heckmann BL, Green DR. The clearance of dead cells by efferocytosis. Nat Rev Mol Cell Biol. 2020;21:398–414.
Rothlin CV, Hille TD, Ghosh S. Determining the effector response to cell death. Nat Rev Immunol. 2021;21:292–304.
Morioka S, Maueroder C, Ravichandran KS. Living on the edge: efferocytosis at the interface of homeostasis and pathology. Immunity. 2019;50:1149–62.
Raymond MH, Davidson AJ, Shen Y, Tudor DR, Lucas CD, Morioka S, et al. Live cell tracking of macrophage efferocytosis during Drosophila embryo development in vivo. Science. 2022;375:1182–7.
Nonomura K, Yamaguchi Y, Hamachi M, Koike M, Uchiyama Y, Nakazato K, et al. Local apoptosis modulates early mammalian brain development through the elimination of morphogen-producing cells. Dev Cell. 2013;27:621–34.
Li MO, Sarkisian MR, Mehal WZ, Rakic P, Flavell RA. Phosphatidylserine receptor is required for clearance of apoptotic cells. Science. 2003;302:1560–3.
Kist M, Vucic D. Cell death pathways: intricate connections and disease implications. EMBO J. 2021;40:e106700.
Doerflinger M, Deng Y, Whitney P, Salvamoser R, Engel S, Kueh AJ, et al. Flexible usage and interconnectivity of diverse cell death pathways protect against intracellular infection. Immunity. 2020;53:533–47.e537.
Oberst A, Dillon CP, Weinlich R, McCormick LL, Fitzgerald P, Pop C, et al. Catalytic activity of the caspase-8-FLIP(L) complex inhibits RIPK3-dependent necrosis. Nature. 2011;471:363–7.
Kaiser WJ, Upton JW, Long AB, Livingston-Rosanoff D, Daley-Bauer LP, Hakem R, et al. RIP3 mediates the embryonic lethality of caspase-8-deficient mice. Nature. 2011;471:368–72.
O’Donnell MA, Perez-Jimenez E, Oberst A, Ng A, Massoumi R, Xavier R, et al. Caspase 8 inhibits programmed necrosis by processing CYLD. Nat Cell Biol. 2011;13:1437–42.
Cauwels A, Janssen B, Waeytens A, Cuvelier C, Brouckaert P. Caspase inhibition causes hyperacute tumor necrosis factor-induced shock via oxidative stress and phospholipase A2. Nat Immunol. 2003;4:387–93.
Vercammen D, Brouckaert G, Denecker G, Van de Craen M, Declercq W, Fiers W, et al. Dual signaling of the Fas receptor: initiation of both apoptotic and necrotic cell death pathways. J Exp Med. 1998;188:919–30.
Vercammen D, Beyaert R, Denecker G, Goossens V, Van Loo G, Declercq W, et al. Inhibition of caspases increases the sensitivity of L929 cells to necrosis mediated by tumor necrosis factor. J Exp Med. 1998;187:1477–85.
Brumatti G, Ma C, Lalaoui N, Nguyen NY, Navarro M, Tanzer MC, et al. The caspase-8 inhibitor emricasan combines with the SMAC mimetic birinapant to induce necroptosis and treat acute myeloid leukemia. Sci Transl Med. 2016;8:339ra369.
Saelens X, Kalai M, Vandenabeele P. Translation inhibition in apoptosis: caspase-dependent PKR activation and eIF2-alpha phosphorylation. J Biol Chem. 2001;276:41620–8.
Tait SW, Green DR. Mitochondria and cell death: outer membrane permeabilization and beyond. Nat Rev Mol Cell Biol. 2010;11:621–32.
Chipuk JE, Mohammed JN, Gelles JD, Chen Y. Mechanistic connections between mitochondrial biology and regulated cell death. Dev Cell. 2021;56:1221–33.
Green DR, Victor B. The pantheon of the fallen: why are there so many forms of cell death? Trends Cell Biol. 2012;22:555–6.
Davidovich P, Kearney CJ, Martin SJ. Inflammatory outcomes of apoptosis, necrosis and necroptosis. Biol Chem. 2014;395:1163–71.
Galluzzi L, López-Soto A, Kumar S, Kroemer G. Caspases connect cell-death signaling to organismal homeostasis. Immunity. 2016;44:221–31.
Duprez L, Takahashi N, Van Hauwermeiren F, Vandendriessche B, Goossens V, Vanden Berghe T, et al. RIP kinase-dependent necrosis drives lethal systemic inflammatory response syndrome. Immunity. 2011;35:908–18.
Glab JA, Cao Z, Puthalakath H. Bcl-2 family proteins, beyond the veil. Int Rev Cell Mol Biol. 2020;351:1–22.
Gross A, Katz SG. Non-apoptotic functions of BCL-2 family proteins. Cell Death Differ. 2017;24:1348–58.
Hollville E, Deshmukh M. Physiological functions of non-apoptotic caspase activity in the nervous system. Semin Cell Dev Biol. 2018;82:127–36.
Nakajima YI, Kuranaga E. Caspase-dependent non-apoptotic processes in development. Cell Death Differ. 2017;24:1422–30.
Aram L, Yacobi-Sharon K, Arama E. CDPs: caspase-dependent non-lethal cellular processes. Cell Death Differ. 2017;24:1307–10.
Feinstein-Rotkopf Y, Arama E. Can’t live without them, can live with them: roles of caspases during vital cellular processes. Apoptosis. 2009;14:980–95.
Perciavalle RM, Stewart DP, Koss B, Lynch J, Milasta S, Bathina M, et al. Anti-apoptotic MCL-1 localizes to the mitochondrial matrix and couples mitochondrial fusion to respiration. Nat Cell Biol. 2012;14:575–83.
Wu L, Tan JL, Wang ZH, Chen YX, Gao L, Liu JL, et al. ROS generated during early reperfusion contribute to intermittent hypobaric hypoxia-afforded cardioprotection against postischemia-induced Ca(2+) overload and contractile dysfunction via the JAK2/STAT3 pathway. J Mol Cell Cardiol. 2015;81:150–61.
Vanden Berghe T, Hulpiau P, Martens L, Vandenbroucke RE, Van Wonterghem E, Perry SW, et al. Passenger mutations confound interpretation of all genetically modified congenic mice. Immunity. 2015;43:200–9.
Wang Y, Gao W, Shi X, Ding J, Liu W, He H, et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature. 2017;547:99–103.
Rogers C, Fernandes-Alnemri T, Mayes L, Alnemri D, Cingolani G, Alnemri ES. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat Commun. 2017;8:14128.
Gould TW, Buss RR, Vinsant S, Prevette D, Sun W, Knudson CM, et al. Complete dissociation of motor neuron death from motor dysfunction by Bax deletion in a mouse model of ALS. J Neurosci. 2006;26:8774–86.
Reyes NA, Fisher JK, Austgen K, VandenBerg S, Huang EJ, Oakes SA. Blocking the mitochondrial apoptotic pathway preserves motor neuron viability and function in a mouse model of amyotrophic lateral sclerosis. J Clin Investig. 2010;120:3673–9.
Kostic V, Jackson-Lewis V, Dubois-Dauphin M, Przedborski S. Bcl-2: prolonging life in a transgenic mouse model of familial amyotrophic lateral sclerosis. Science. 1997;277:559–62.
Vukosavic S, Stefanis L, Jackson-Lewis V, Guégan C, Romero N, Chen C, et al. Delaying caspase activation by Bcl-2: A clue to disease retardation in a transgenic mouse model of amyotrophic lateral sclerosis. J Neurosci. 2000;20:9119–25.
Inoue H, Tsukita K, Iwasato T, Suzuki Y, Tomioka M, Tateno M, et al. The crucial role of caspase-9 in the disease progression of a transgenic ALS mouse model. EMBO J. 2003;22:6665–74.
Wootz H, Hansson I, Korhonen L, Lindholm D. XIAP decreases caspase-12 cleavage and calpain activity in spinal cord of ALS transgenic mice. Exp Cell Res. 2006;312:1890–8.
Kieran D, Woods I, Villunger A, Strasser A, Prehn JH. Deletion of the BH3-only protein puma protects motoneurons from ER stress-induced apoptosis and delays motoneuron loss in ALS mice. Proc Natl Acad Sci USA. 2007;104:20606–11.
Li M, Ona VO, Guégan C, Chen M, Jackson-Lewis V, Andrews LJ, et al. Functional role of caspase-1 and caspase-3 in an ALS transgenic mouse model. Science. 2000;288:335–9.
Girgenrath M, Dominov JA, Kostek CA, Miller JB. Inhibition of apoptosis improves outcome in a model of congenital muscular dystrophy. J Clin Investig. 2004;114:1635–9.
Davies JE, Rubinsztein DC. Over-expression of BCL2 rescues muscle weakness in a mouse model of oculopharyngeal muscular dystrophy. Hum Mol Genet. 2011;20:1154–63.
Dominov JA, Kravetz AJ, Ardelt M, Kostek CA, Beermann ML, Miller JB. Muscle-specific BCL2 expression ameliorates muscle disease in laminin {alpha}2-deficient, but not in dystrophin-deficient, mice. Hum Mol Genet. 2005;14:1029–40.
Sagot Y, Dubois-Dauphin M, Tan SA, de Bilbao F, Aebischer P, Martinou JC, et al. Bcl-2 overexpression prevents motoneuron cell body loss but not axonal degeneration in a mouse model of a neurodegenerative disease. J Neurosci. 1995;15:7727–33.
Tossing G, Livernoche R, Maios C, Bretonneau C, Labarre A, Parker JA. Genetic and pharmacological PARP inhibition reduces axonal degeneration in C. elegans models of ALS. Hum Mol Genet. 2022;31:3313–24.
Kudo W, Lee HP, Smith MA, Zhu X, Matsuyama S, Lee HG. Inhibition of Bax protects neuronal cells from oligomeric Aβ neurotoxicity. Cell Death Dis. 2012;3:e309.
Bové J, Martínez-Vicente M, Dehay B, Perier C, Recasens A, Bombrun A, et al. BAX channel activity mediates lysosomal disruption linked to Parkinson disease. Autophagy. 2014;10:889–900.
Vila M, Jackson-Lewis V, Vukosavic S, Djaldetti R, Liberatore G, Offen D, et al. Bax ablation prevents dopaminergic neurodegeneration in the 1-methyl- 4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. Proc Natl Acad Sci USA. 2001;98:2837–42.
Kim TW, Moon Y, Kim K, Lee JE, Koh HC, Rhyu IJ, et al. Dissociation of progressive dopaminergic neuronal death and behavioral impairments by Bax deletion in a mouse model of Parkinson’s diseases. PloS one. 2011;6:e25346.
Ma C, Pan Y, Yang Z, Meng Z, Sun R, Wang T, et al. Pre-administration of BAX-inhibiting peptides decrease the loss of the nigral dopaminergic neurons in rats. Life Sci. 2016;144:113–20.
Jiang H, He P, Adler CH, Shill H, Beach TG, Li R, et al. Bid signal pathway components are identified in the temporal cortex with Parkinson disease. Neurology. 2012;79:1767–73.
Biswas SC, Ryu E, Park C, Malagelada C, Greene LA. Puma and p53 play required roles in death evoked in a cellular model of Parkinson disease. Neurochem Res. 2005;30:839–45.
Akhter R, Saleem S, Saha A, Biswas SC. The pro-apoptotic protein Bmf co-operates with Bim and Puma in neuron death induced by β-amyloid or NGF deprivation. Mol Cell Neurosci. 2018;88:249–57.
Imaizumi K, Morihara T, Mori Y, Katayama T, Tsuda M, Furuyama T, et al. The cell death-promoting gene DP5, which interacts with the BCL2 family, is induced during neuronal apoptosis following exposure to amyloid beta protein. J Biol Chem. 1999;274:7975–81.
Louneva N, Cohen JW, Han LY, Talbot K, Wilson RS, Bennett DA, et al. Caspase-3 is enriched in postsynaptic densities and increased in Alzheimer’s disease. Am J Pathol. 2008;173:1488–95.
Rohn TT, Rissman RA, Davis MC, Kim YE, Cotman CW, Head E. Caspase-9 activation and caspase cleavage of tau in the Alzheimer’s disease brain. Neurobiol Dis. 2002;11:341–54.
Hartmann A, Hunot S, Michel PP, Muriel MP, Vyas S, Faucheux BA, et al. Caspase-3: a vulnerability factor and final effector in apoptotic death of dopaminergic neurons in Parkinson’s disease. Proc Natl Acad Sci USA. 2000;97:2875–80.
Zhang L, Qian Y, Li J, Zhou X, Xu H, Yan J, et al. BAD-mediated neuronal apoptosis and neuroinflammation contribute to Alzheimer’s disease pathology. iScience. 2021;24:102942.
Rissman RA, Poon WW, Blurton-Jones M, Oddo S, Torp R, Vitek MP, et al. Caspase-cleavage of tau is an early event in Alzheimer disease tangle pathology. J Clin Investig. 2004;114:121–30.
Gervais FG, Xu D, Robertson GS, Vaillancourt JP, Zhu Y, Huang J, et al. Involvement of caspases in proteolytic cleavage of Alzheimer’s amyloid-beta precursor protein and amyloidogenic A beta peptide formation. Cell. 1999;97:395–406.
Chu J, Lauretti E, Praticò D. Caspase-3-dependent cleavage of Akt modulates tau phosphorylation via GSK3β kinase: implications for Alzheimer’s disease. Mol Psychiatry. 2017;22:1002–8.
Rohn TT, Vyas V, Hernandez-Estrada T, Nichol KE, Christie LA, Head E. Lack of pathology in a triple transgenic mouse model of Alzheimer’s disease after overexpression of the anti-apoptotic protein Bcl-2. J Neurosci. 2008;28:3051–9.
Kumasaka DK, Galvan V, Head E, Rohn TT. Caspase cleavage of the amyloid precursor protein is prevented after overexpression of bcl-2 in a triple transgenic mouse model of Alzheimer’s disease. Int J Physiol, Pathophysiol Pharmacol. 2009;1:48–56.
D’Amelio M, Cavallucci V, Middei S, Marchetti C, Pacioni S, Ferri A, et al. Caspase-3 triggers early synaptic dysfunction in a mouse model of Alzheimer’s disease. Nat Neurosci. 2011;14:69–76.
Park G, Nhan HS, Tyan SH, Kawakatsu Y, Zhang C, Navarro M, et al. Caspase activation and caspase-mediated cleavage of APP is associated with amyloid β-protein-induced synapse loss in Alzheimer’s disease. Cell Rep. 2020;31:107839.
Pozueta J, Lefort R, Ribe EM, Troy CM, Arancio O, Shelanski M. Caspase-2 is required for dendritic spine and behavioural alterations in J20 APP transgenic mice. Nat Commun. 2013;4:1939.
Troy CM, Shelanski ML. Caspase-2 and tau-a toxic partnership? Nat Med. 2016;22:1207–8.
Zhao X, Kotilinek LA, Smith B, Hlynialuk C, Zahs K, Ramsden M, et al. Caspase-2 cleavage of tau reversibly impairs memory. Nat Med. 2016;22:1268–76.
Steuer EL, Kemper LJ, Hlynialuk CJW, Leinonen-Wright K, Montonye ML, Lapcinski IP, et al. Blocking site-specific cleavage of human tau delays progression of disease-related phenotypes in genetically matched tau-transgenic mice modeling frontotemporal dementia. J Neurosci. 2022;42:4737–54.
Bresinsky M, Strasser JM, Vallaster B, Liu P, McCue WM, Fuller J, et al. Structure-based design and biological evaluation of novel caspase-2 inhibitors based on the peptide AcVDVAD-CHO and the caspase-2-mediated tau cleavage sequence YKPVD314. ACS Pharmacol Transl Sci. 2022;5:20–40.
Kajiwara Y, McKenzie A, Dorr N, Gama Sosa MA, Elder G, Schmeidler J, et al. The human-specific CASP4 gene product contributes to Alzheimer-related synaptic and behavioural deficits. Hum Mol Genet. 2016;25:4315–27.
Lee JH, Won SM, Suh J, Son SJ, Moon GJ, Park UJ, et al. Induction of the unfolded protein response and cell death pathway in Alzheimer’s disease, but not in aged Tg2576 mice. Exp Mol Med. 2010;42:386–94.
Kolosova NG, Tyumentsev MA, Muraleva NA, Kiseleva E, Vitovtov AO, Stefanova NA. Antioxidant SkQ1 alleviates signs of Alzheimer’s disease-like pathology in old OXYS rats by reversing mitochondrial deterioration. Curr Alzheimer Res. 2017;14:1283–92.
Perier C, Bové J, Wu DC, Dehay B, Choi DK, Jackson-Lewis V, et al. Two molecular pathways initiate mitochondria-dependent dopaminergic neurodegeneration in experimental Parkinson’s disease. Proc Natl Acad Sci USA. 2007;104:8161–6.
Yamada M, Kida K, Amutuhaire W, Ichinose F, Kaneki M. Gene disruption of caspase-3 prevents MPTP-induced Parkinson’s disease in mice. Biochem Biophys Res Commun. 2010;402:312–8.
Viswanath V, Wu Y, Boonplueang R, Chen S, Stevenson FF, Yantiri F, et al. Caspase-9 activation results in downstream caspase-8 activation and bid cleavage in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson’s disease. J Neurosci. 2001;21:9519–28.
Crocker SJ, Liston P, Anisman H, Lee CJ, Smith PD, Earl N, et al. Attenuation of MPTP-induced neurotoxicity and behavioural impairment in NSE-XIAP transgenic mice. Neurobiol Dis. 2003;12:150–61.
Liu Y, Guo Y, An S, Kuang Y, He X, Ma H, et al. Targeting caspase-3 as dual therapeutic benefits by RNAi facilitating brain-targeted nanoparticles in a rat model of Parkinson’s disease. PLoS ONE. 2013;8:e62905.
Toulmond S, Tang K, Bureau Y, Ashdown H, Degen S, O’Donnell R, et al. Neuroprotective effects of M826, a reversible caspase-3 inhibitor, in the rat malonate model of Huntington’s disease. Br J Pharmacol. 2004;141:689–97.
Leyva MJ, Degiacomo F, Kaltenbach LS, Holcomb J, Zhang N, Gafni J, et al. Identification and evaluation of small molecule pan-caspase inhibitors in Huntington’s disease models. Chem Biol. 2010;17:1189–1200.
Chen M, Ona VO, Li M, Ferrante RJ, Fink KB, Zhu S, et al. Minocycline inhibits caspase-1 and caspase-3 expression and delays mortality in a transgenic mouse model of Huntington disease. Nat Med. 2000;6:797–801.
Cen X, Chen Y, Xu X, Wu R, He F, Zhao Q, et al. Pharmacological targeting of MCL-1 promotes mitophagy and improves disease pathologies in an Alzheimer’s disease mouse model. Nat Commun. 2020;11:5731.
Ekholm-Reed S, Baker R, Campos AR, Stouffer D, Henze M, Wolf DA, et al. Reducing Mcl-1 gene dosage induces dopaminergic neuronal loss and motor impairments in Park2 knockout mice. Commun Biol. 2019;2:125.
Koper MJ, Van Schoor E, Ospitalieri S, Vandenberghe R, Vandenbulcke M, von Arnim CAF, et al. Necrosome complex detected in granulovacuolar degeneration is associated with neuronal loss in Alzheimer’s disease. Acta Neuropathol. 2020;139:463–84.
Hambright WS, Fonseca RS, Chen L, Na R, Ran Q. Ablation of ferroptosis regulator glutathione peroxidase 4 in forebrain neurons promotes cognitive impairment and neurodegeneration. Redox Biol. 2017;12:8–17.
Chiesa R, Piccardo P, Dossena S, Nowoslawski L, Roth KA, Ghetti B, et al. Bax deletion prevents neuronal loss but not neurological symptoms in a transgenic model of inherited prion disease. Proc Natl Acad Sci USA. 2005;102:238–43.
Steele AD, King OD, Jackson WS, Hetz CA, Borkowski AW, Thielen P, et al. Diminishing apoptosis by deletion of Bax or overexpression of Bcl-2 does not protect against infectious prion toxicity in vivo. J Neurosci. 2007;27:13022–7.
Pemberton JM, Pogmore JP, Andrews DW. Neuronal cell life, death, and axonal degeneration as regulated by the BCL-2 family proteins. Cell Death Differ. 2021;28:108–22.
Ray SK, Samantaray S, Smith JA, Matzelle DD, Das A, Banik NL. Inhibition of cysteine proteases in acute and chronic spinal cord injury. Neurotherapeutics. 2011;8:180–6.
Sarosiek KA, Fraser C, Muthalagu N, Bhola PD, Chang W, McBrayer SK, et al. Developmental regulation of mitochondrial apoptosis by c-Myc governs age- and tissue-specific sensitivity to cancer therapeutics. Cancer Cell. 2017;31:142–56.
Tehranian R, Rose ME, Vagni V, Pickrell AM, Griffith RP, Liu H, et al. Disruption of Bax protein prevents neuronal cell death but produces cognitive impairment in mice following traumatic brain injury. J Neurotrauma. 2008;25:755–67.
Tehranian R, Rose ME, Vagni V, Griffith RP, Wu S, Maits S, et al. Transgenic mice that overexpress the anti-apoptotic Bcl-2 protein have improved histological outcome but unchanged behavioral outcome after traumatic brain injury. Brain Res. 2006;1101:126–35.
Bermpohl D, You Z, Korsmeyer SJ, Moskowitz MA, Whalen MJ. Traumatic brain injury in mice deficient in Bid: effects on histopathology and functional outcome. J Cereb Blood Flow Metab. 2006;26:625–33.
Raghupathi R, Fernandez SC, Murai H, Trusko SP, Scott RW, Nishioka WK, et al. BCL-2 overexpression attenuates cortical cell loss after traumatic brain injury in transgenic mice. J Cereb Blood Flow Metab. 1998;18:1259–69.
Farlie PG, Dringen R, Rees SM, Kannourakis G, Bernard O. bcl-2 transgene expression can protect neurons against developmental and induced cell death. Proc Natl Acad Sci USA. 1995;92:4397–401.
Dong H, Fazzaro A, Xiang C, Korsmeyer SJ, Jacquin MF, McDonald JW. Enhanced oligodendrocyte survival after spinal cord injury in Bax-deficient mice and mice with delayed Wallerian degeneration. J Neurosci. 2003;23:8682–91.
Barut S, Unlü YA, Karaoğlan A, Tunçdemir M, Dağistanli FK, Oztürk M, et al. The neuroprotective effects of z-DEVD.fmk, a caspase-3 inhibitor, on traumatic spinal cord injury in rats. Surg Neurol. 2005;64:213–20.
Colak A, Karaoğlan A, Barut S, Köktürk S, Akyildiz AI, Taşyürekli M. Neuroprotection and functional recovery after application of the caspase-9 inhibitor z-LEHD-fmk in a rat model of traumatic spinal cord injury. J Neurosurg Spine. 2005;2:327–34.
Li M, Ona VO, Chen M, Kaul M, Tenneti L, Zhang X, et al. Functional role and therapeutic implications of neuronal caspase-1 and -3 in a mouse model of traumatic spinal cord injury. Neuroscience. 2000;99:333–42.
Zhao W, Li H, Hou Y, Jin Y, Zhang L. Combined administration of poly-ADP-ribose polymerase-1 and caspase-3 inhibitors alleviates neuronal apoptosis after spinal cord injury in rats. World Neurosurg. 2019;127:e346–e352.
Donahue RJ, Maes ME, Grosser JA, Nickells RW. BAX-depleted retinal ganglion cells survive and become quiescent following optic nerve damage. Mol Neurobiol. 2020;57:1070–84.
Libby RT, Li Y, Savinova OV, Barter J, Smith RS, Nickells RW, et al. Susceptibility to neurodegeneration in a glaucoma is modified by Bax gene dosage. PLoS Genet. 2005;1:17–26.
Harder JM, Libby RT. BBC3 (PUMA) regulates developmental apoptosis but not axonal injury induced death in the retina. Mol Neurodegener. 2011;6:50.
Harder JM, Libby RT. Deficiency in Bim, Bid and Bbc3 (Puma) do not prevent axonal injury induced death. Cell Death Differ. 2013;20:182.
Harder JM, Ding Q, Fernandes KA, Cherry JD, Gan L, Libby RT. BCL2L1 (BCL-X) promotes survival of adult and developing retinal ganglion cells. Mol Cell Neurosci. 2012;51:53–9.
Visuvanathan S, Baker AN, Lagali PS, Coupland SG, Miller G, Hauswirth WW, et al. XIAP gene therapy effects on retinal ganglion cell structure and function in a mouse model of glaucoma. Gene Ther. 2022;29:147–56.
Donahue RJ, Fehrman RL, Gustafson JR, Nickells RW. BCLX(L) gene therapy moderates neuropathology in the DBA/2J mouse model of inherited glaucoma. Cell Death Dis. 2021;12:781.
Avrutsky MI, Ortiz CC, Johnson KV, Potenski AM, Chen CW, Lawson JM, et al. Endothelial activation of caspase-9 promotes neurovascular injury in retinal vein occlusion. Nat Commun. 2020;11:3173.
Ishikawa S, Hirata A, Nakabayashi J, Iwakiri R, Okinami S. Neuroprotective effect of small interfering RNA targeted to caspase-3 on rat retinal ganglion cell loss induced by ischemia and reperfusion injury. Curr Eye Res. 2012;37:907–13.
Tawfik M, Zhang X, Grigartzik L, Heiduschka P, Hintz W, Henrich-Noack P, et al. Gene therapy with caspase-3 small interfering RNA-nanoparticles is neuroprotective after optic nerve damage. Neural Regen Res. 2021;16:2534–41.
Wassmer SJ, De Repentigny Y, Sheppard D, Lagali PS, Fang L, Coupland SG, et al. XIAP protects retinal ganglion cells in the mutant ND4 mouse model of leber hereditary optic neuropathy. Investig Ophthalmol Vis Sci. 2020;61:49.
Wassmer SJ, Leonard BC, Coupland SG, Baker AN, Hamilton J, Hauswirth WW, et al. Overexpression of the X-linked inhibitor of apoptosis protects against retinal degeneration in a feline model of retinal detachment. Hum Gene Ther. 2017;28:482–92.
Renwick J, Narang MA, Coupland SG, Xuan JY, Baker AN, Brousseau J, et al. XIAP-mediated neuroprotection in retinal ischemia. Gene Ther. 2006;13:339–47.
McKinnon SJ, Lehman DM, Tahzib NG, Ransom NL, Reitsamer HA, Liston P, et al. Baculoviral IAP repeat-containing-4 protects optic nerve axons in a rat glaucoma model. Mol Ther. 2002;5:780–7.
Zadro-Lamoureux LA, Zacks DN, Baker AN, Zheng QD, Hauswirth WW, Tsilfidis C. XIAP effects on retinal detachment-induced photoreceptor apoptosis [corrected]. Investig Ophthalmol Vis Sci. 2009;50:1448–53.
Yao J, Feathers KL, Khanna H, Thompson D, Tsilfidis C, Hauswirth WW, et al. XIAP therapy increases survival of transplanted rod precursors in a degenerating host retina. Investig Ophthalmol Vis Sci. 2011;52:1567–72.
Crespo-Garcia S, Tsuruda PR, Dejda A, Ryan RD, Fournier F, Chaney SY, et al. Pathological angiogenesis in retinopathy engages cellular senescence and is amenable to therapeutic elimination via BCL-xL inhibition. Cell Metab. 2021;33:818–32.e817.
Choudhury S, Liu Y, Clark AF, Pang IH. Caspase-7: a critical mediator of optic nerve injury-induced retinal ganglion cell death. Mol Neurodegener. 2015;10:40.
Wang S, Sorenson CM, Sheibani N. Attenuation of retinal vascular development and neovascularization during oxygen-induced ischemic retinopathy in Bcl-2−/− mice. Dev Biol. 2005;279:205–19.
Wang S, Park S, Fei P, Sorenson CM. Bim is responsible for the inherent sensitivity of the developing retinal vasculature to hyperoxia. Dev Biol. 2011;349:296–309.
Grant ZL, Whitehead L, Wong VH, He Z, Yan RY, Miles AR, et al. Blocking endothelial apoptosis revascularizes the retina in a model of ischemic retinopathy. J Clin Investig. 2020;130:4235–51.
Du H, Sun X, Guma M, Luo J, Ouyang H, Zhang X, et al. JNK inhibition reduces apoptosis and neovascularization in a murine model of age-related macular degeneration. Proc Natl Acad Sci USA. 2013;110:2377–82.
Deckwerth TL, Elliott JL, Knudson CM, Johnson EM Jr, Snider WD, Korsmeyer SJ. BAX is required for neuronal death after trophic factor deprivation and during development. Neuron. 1996;17:401–11.
Unsain N, Higgins JM, Parker KN, Johnstone AD, Barker PA. XIAP regulates caspase activity in degenerating axons. Cell Rep. 2013;4:751–63.
Imaizumi K, Benito A, Kiryu-Seo S, Gonzalez V, Inohara N, Lieberman AP, et al. Critical role for DP5/Harakiri, a Bcl-2 homology domain 3-only Bcl-2 family member, in axotomy-induced neuronal cell death. J Neurosci. 2004;24:3721–5.
Theofilas P, Bedner P, Hüttmann K, Theis M, Steinhäuser C, Frank S. The proapoptotic BCL-2 homology domain 3-only protein Bim is not critical for acute excitotoxic cell death. J Neuropathol Exp Neurol. 2009;68:102–10.
Bunk EC, König HG, Prehn JHM, Kirby BP. p53 upregulated mediator of apoptosis (Puma) deficiency increases survival of adult neural stem cells generated physiologically in the hippocampus, but does not protect stem cells generated in surplus after an excitotoxic lesion. J Basic Clin Physiol Pharmacol. 2020;32:57–66.
Li T, Fan Y, Luo Y, Xiao B, Lu C. In vivo delivery of a XIAP (BIR3-RING) fusion protein containing the protein transduction domain protects against neuronal death induced by seizures. Exp Neurol. 2006;197:301–8.
Tzeng TT, Tsay HJ, Chang L, Hsu CL, Lai TH, Huang FL, et al. Caspase 3 involves in neuroplasticity, microglial activation and neurogenesis in the mice hippocampus after intracerebral injection of kainic acid. J Biomed Sci. 2013;20:90.
Concannon CG, Tuffy LP, Weisová P, Bonner HP, Dávila D, Bonner C, et al. AMP kinase-mediated activation of the BH3-only protein Bim couples energy depletion to stress-induced apoptosis. J Cell Biol. 2010;189:83–94.
Murphy BM, Engel T, Paucard A, Hatazaki S, Mouri G, Tanaka K, et al. Contrasting patterns of Bim induction and neuroprotection in Bim-deficient mice between hippocampus and neocortex after status epilepticus. Cell Death Differ. 2010;17:459–68.
Foley J, Burnham V, Tedoldi M, Danial NN, Yellen G. BAD knockout provides metabolic seizure resistance in a genetic model of epilepsy with sudden unexplained death in epilepsy. Epilepsia. 2018;59:e1–e4.
Moran C, Sanz-Rodriguez A, Jimenez-Pacheco A, Martinez-Villareal J, McKiernan RC, Jimenez-Mateos EM, et al. Bmf upregulation through the AMP-activated protein kinase pathway may protect the brain from seizure-induced cell death. Cell Death Dis. 2013;4:e606.
Engel T, Murphy BM, Hatazaki S, Jimenez-Mateos EM, Concannon CG, Woods I, et al. Reduced hippocampal damage and epileptic seizures after status epilepticus in mice lacking proapoptotic Puma. FASEB J. 2010;24:853–61.
Engel T, Hatazaki S, Tanaka K, Prehn JH, Henshall DC. Deletion of Puma protects hippocampal neurons in a model of severe status epilepticus. Neuroscience. 2010;168:443–50.
Murphy B, Dunleavy M, Shinoda S, Schindler C, Meller R, Bellver-Estelles C, et al. Bcl-w protects hippocampus during experimental status epilepticus. Am J Pathol. 2007;171:1258–68.
Ichikawa N, Alves M, Pfeiffer S, Langa E, Hernández-Santana YE, Suzuki H, et al. Deletion of the BH3-only protein Noxa alters electrographic seizures but does not protect against hippocampal damage after status epilepticus in mice. Cell Death Dis. 2017;8:e2556.
Engel T, Caballero-Caballero A, Schindler CK, Plesnila N, Strasser A, Prehn JH, et al. BH3-only protein Bid is dispensable for seizure-induced neuronal death and the associated nuclear accumulation of apoptosis-inducing factor. J Neurochem. 2010;115:92–101.
Gibson ME, Han BH, Choi J, Knudson CM, Korsmeyer SJ, Parsadanian M, et al. BAX contributes to apoptotic-like death following neonatal hypoxia-ischemia: evidence for distinct apoptosis pathways. Mol Med. 2001;7:644–55.
Ness JM, Harvey CA, Strasser A, Bouillet P, Klocke BJ, Roth KA. Selective involvement of BH3-only Bcl-2 family members Bim and Bad in neonatal hypoxia-ischemia. Brain Res. 2006;1099:150–9.
Wang X, Zhu C, Wang X, Hagberg H, Korhonen L, Sandberg M, et al. X-linked inhibitor of apoptosis (XIAP) protein protects against caspase activation and tissue loss after neonatal hypoxia-ischemia. Neurobiol Dis. 2004;16:179–89.
West T, Stump M, Lodygensky G, Neil JJ, Deshmukh M, Holtzman DM. Lack of X-linked inhibitor of apoptosis protein leads to increased apoptosis and tissue loss following neonatal brain injury. ASN Neuro 2009;1:e00004.
West T, Atzeva M, Holtzman DM. Caspase-3 deficiency during development increases vulnerability to hypoxic-ischemic injury through caspase-3-independent pathways. Neurobiol Dis. 2006;22:523–37.
Ghosh AP, Walls KC, Klocke BJ, Toms R, Strasser A, Roth KA. The proapoptotic BH3-only, Bcl-2 family member, Puma is critical for acute ethanol-induced neuronal apoptosis. J Neuropathol Exp Neurol. 2009;68:747–56.
Young C, Klocke BJ, Tenkova T, Choi J, Labruyere J, Qin YQ, et al. Ethanol-induced neuronal apoptosis in vivo requires BAX in the developing mouse brain. Cell Death Differ. 2003;10:1148–55.
Young C, Roth KA, Klocke BJ, West T, Holtzman DM, Labruyere J, et al. Role of caspase-3 in ethanol-induced developmental neurodegeneration. Neurobiol Dis. 2005;20:608–14.
Slupe AM, Villasana L, Wright KM. GABAergic neurons are susceptible to BAX-dependent apoptosis following isoflurane exposure in the neonatal period. PLoS ONE. 2021;16:e0238799.
Chong MJ, Murray MR, Gosink EC, Russell HR, Srinivasan A, Kapsetaki M, et al. Atm and Bax cooperate in ionizing radiation-induced apoptosis in the central nervous system. Proc Natl Acad Sci USA. 2000;97:889–94.
Ahlers KE, Karaçay B, Fuller L, Bonthius DJ, Dailey ME. Transient activation of microglia following acute alcohol exposure in developing mouse neocortex is primarily driven by BAX-dependent neurodegeneration. Glia. 2015;63:1694–713.
D’Orsi B, Kilbride SM, Chen G, Perez Alvarez S, Bonner HP, Pfeiffer S, et al. Bax regulates neuronal Ca2+ homeostasis. J Neurosci. 2015;35:1706–22.
Pfeiffer S, Anilkumar U, Chen G, Ramírez-Peinado S, Galindo-Moreno J, Muñoz-Pinedo C, et al. Analysis of BH3-only proteins upregulated in response to oxygen/glucose deprivation in cortical neurons identifies Bmf but not Noxa as potential mediator of neuronal injury. Cell Death Dis. 2014;5:e1456.
Plesnila N, Zinkel S, Le DA, Amin-Hanjani S, Wu Y, Qiu J, et al. BID mediates neuronal cell death after oxygen/ glucose deprivation and focal cerebral ischemia. Proc Natl Acad Sci USA. 2001;98:15318–23.
Yin XM, Luo Y, Cao G, Bai L, Pei W, Kuharsky DK, et al. Bid-mediated mitochondrial pathway is critical to ischemic neuronal apoptosis and focal cerebral ischemia. J Biol Chem. 2002;277:42074–81.
Plesnila N, Zinkel S, Amin-Hanjani S, Qiu J, Korsmeyer SJ, Moskowitz MA. Function of BID - a molecule of the bcl-2 family - in ischemic cell death in the brain. Eur Surg Res. 2002;34:37–41.
Martin NA, Bonner H, Elkjær ML, D’Orsi B, Chen G, König HG, et al. BID mediates oxygen-glucose deprivation-induced neuronal injury in organotypic hippocampal slice cultures and modulates tissue inflammation in a transient focal cerebral ischemia model without changing lesion volume. Front Cell Neurosci. 2016;10:14.
Kitagawa K, Matsumoto M, Tsujimoto Y, Ohtsuki T, Kuwabara K, Matsushita K, et al. Amelioration of hippocampal neuronal damage after global ischemia by neuronal overexpression of BCL-2 in transgenic mice. Stroke. 1998;29:2616–21.
Cao G, Pei W, Ge H, Liang Q, Luo Y, Sharp FR, et al. In vivo delivery of a Bcl-xL fusion protein containing the TAT protein transduction domain protects against ischemic brain injury and neuronal apoptosis. J Neurosci. 2002;22:5423–31.
Kilic E, Hermann DM, Kügler S, Kilic U, Holzmüller H, Schmeer C, et al. Adenovirus-mediated Bcl-X(L) expression using a neuron-specific synapsin-1 promoter protects against disseminated neuronal injury and brain infarction following focal cerebral ischemia in mice. Neurobiol Dis. 2002;11:275–84.
Akpan N, Serrano-Saiz E, Zacharia BE, Otten ML, Ducruet AF, Snipas SJ, et al. Intranasal delivery of caspase-9 inhibitor reduces caspase-6-dependent axon/neuron loss and improves neurological function after stroke. J Neurosci. 2011;31:8894–904.
Fan YF, Lu CZ, Xie J, Zhao YX, Yang GY. Apoptosis inhibition in ischemic brain by intraperitoneal PTD-BIR3-RING (XIAP). Neurochem Int. 2006;48:50–59.
Trapp T, Korhonen L, Besselmann M, Martinez R, Mercer EA, Lindholm D. Transgenic mice overexpressing XIAP in neurons show better outcome after transient cerebral ischemia. Mol Cell Neurosci. 2003;23:302–13.
Zhu C, Xu F, Fukuda A, Wang X, Fukuda H, Korhonen L, et al. X chromosome-linked inhibitor of apoptosis protein reduces oxidative stress after cerebral irradiation or hypoxia-ischemia through up-regulation of mitochondrial antioxidants. Eur J Neurosci. 2007;26:3402–10.
Zhao H, Yenari MA, Cheng D, Sapolsky RM, Steinberg GK. Biphasic cytochrome c release after transient global ischemia and its inhibition by hypothermia. J Cereb Blood Flow Metab. 2005;25:1119–29.
Chen J, Nagayama T, Jin K, Stetler RA, Zhu RL, Graham SH, et al. Induction of caspase-3-like protease may mediate delayed neuronal death in the hippocampus after transient cerebral ischemia. J Neurosci. 1998;18:4914–28.
Gao Y, Liang W, Hu X, Zhang W, Stetler RA, Vosler P, et al. Neuroprotection against hypoxic-ischemic brain injury by inhibiting the apoptotic protease activating factor-1 pathway. Stroke. 2010;41:166–72.
Karatas H, Aktas Y, Gursoy-Ozdemir Y, Bodur E, Yemisci M, Caban S, et al. A nanomedicine transports a peptide caspase-3 inhibitor across the blood-brain barrier and provides neuroprotection. J Neurosci. 2009;29:13761–9.
Endres M, Namura S, Shimizu-Sasamata M, Waeber C, Zhang L, Gómez-Isla T, et al. Attenuation of delayed neuronal death after mild focal ischemia in mice by inhibition of the caspase family. J Cereb Blood Flow Metab. 1998;18:238–47.
Gottron FJ, Ying HS, Choi DW. Caspase inhibition selectively reduces the apoptotic component of oxygen-glucose deprivation-induced cortical neuronal cell death. Mol Cell Neurosci. 1997;9:159–69.
Shibata M, Hisahara S, Hara H, Yamawaki T, Fukuuchi Y, Yuan J, et al. Caspases determine the vulnerability of oligodendrocytes in the ischemic brain. J Clin Investig. 2000;106:643–53.
Sung JH, Zhao H, Roy M, Sapolsky RM, Steinberg GK. Viral caspase inhibitor p35, but not crmA, is neuroprotective in the ischemic penumbra following experimental stroke. Neuroscience. 2007;149:804–12.
Braun JS, Prass K, Dirnagl U, Meisel A, Meisel C. Protection from brain damage and bacterial infection in murine stroke by the novel caspase-inhibitor Q-VD-OPH. Exp Neurol. 2007;206:183–91.
Sun Y, Xu Y, Geng L. Caspase-3 inhibitor prevents the apoptosis of brain tissue in rats with acute cerebral infarction. Exp Ther Med. 2015;10:133–8.
Lapchak PA, Araujo DM, Weir CJ, Wei J, Zivin JA. Effects of intrathecal administration of a cell permeant caspase inhibitor, boc-D-fluoromethylketone (BDFMK), on behavioral deficits following spinal cord ischemia: a dose-response analysis. Brain Res. 2003;959:183–90.
Osman AM, Neumann S, Kuhn HG, Blomgren K. Caspase inhibition impaired the neural stem/progenitor cell response after cortical ischemia in mice. Oncotarget. 2016;7:2239–48.
Zhan RZ, Wu C, Fujihara H, Taga K, Qi S, Naito M, et al. Both caspase-dependent and caspase-independent pathways may be involved in hippocampal CA1 neuronal death because of loss of cytochrome c from mitochondria in a rat forebrain ischemia model. J Cereb Blood Flow Metab. 2001;21:529–40.
Moujalled D, Strasser A, Liddell JR. Molecular mechanisms of cell death in neurological diseases. Cell Death Differ. 2021;28:2029–44.
Crowther AJ, Gama V, Bevilacqua A, Chang SX, Yuan H, Deshmukh M, et al. Tonic activation of Bax primes neural progenitors for rapid apoptosis through a mechanism preserved in medulloblastoma. J Neurosci. 2013;33:18098–108.
Nakaya K, Hasegawa T, Flickinger JC, Kondziolka DS, Fellows-Mayle W, Gobbel GT. Sensitivity to radiation-induced apoptosis and neuron loss declines rapidly in the postnatal mouse neocortex. Int J Radiat Biol. 2005;81:545–54.
Kole AJ, Annis RP, Deshmukh M. Mature neurons: equipped for survival. Cell Death Dis. 2013;4:e689.
Whelan RS, Konstantinidis K, Wei AC, Chen Y, Reyna DE, Jha S, et al. Bax regulates primary necrosis through mitochondrial dynamics. Proc Natl Acad Sci USA. 2012;109:6566–71.
Karch J, Kwong JQ, Burr AR, Sargent MA, Elrod JW, Peixoto PM, et al. Bax and Bak function as the outer membrane component of the mitochondrial permeability pore in regulating necrotic cell death in mice. eLife. 2013;2:e00772.
Hochhauser E, Cheporko Y, Yasovich N, Pinchas L, Offen D, Barhum Y, et al. Bax deficiency reduces infarct size and improves long-term function after myocardial infarction. Cell Biochem Biophys. 2007;47:11–20.
Vaseva AV, Marchenko ND, Ji K, Tsirka SE, Holzmann S, Moll UM. p53 opens the mitochondrial permeability transition pore to trigger necrosis. Cell. 2012;149:1536–48.
Brocheriou V, Hagège AA, Oubenaïssa A, Lambert M, Mallet VO, Duriez M, et al. Cardiac functional improvement by a human Bcl-2 transgene in a mouse model of ischemia/reperfusion injury. J Gene Med. 2000;2:326–33.
Kristen AV, Ackermann K, Buss S, Lehmann L, Schnabel PA, Haunstetter A, et al. Inhibition of apoptosis by the intrinsic but not the extrinsic apoptotic pathway in myocardial ischemia-reperfusion. Cardiovasc Pathol. 2013;22:280–6.
Chen Z, Chua CC, Ho YS, Hamdy RC, Chua BH. Overexpression of Bcl-2 attenuates apoptosis and protects against myocardial I/R injury in transgenic mice. Am J Physiol Heart Circ Physiol. 2001;280:H2313–20.
Ono M, Sawa Y, Ryugo M, Alechine AN, Shimizu S, Sugioka R, et al. BH4 peptide derivative from Bcl-xL attenuates ischemia/reperfusion injury thorough anti-apoptotic mechanism in rat hearts. Eur J Cardiothorac Surg. 2005;27:117–21.
Toth A, Jeffers JR, Nickson P, Min JY, Morgan JP, Zambetti GP, et al. Targeted deletion of Puma attenuates cardiomyocyte death and improves cardiac function during ischemia-reperfusion. Am J Physiol Heart Circ Physiol. 2006;291:H52–60.
Gao J, Zhang L, Wang WL, Ma Q, Chu HC. Post-conditioning anti-PUMA treatment protects mice against mice heart I/R injury. Eur Rev Med Pharmacol Sci. 2016;20:1623–7.
Bi W, Wang J, Jiang Y, Li Q, Wang S, Liu M, et al. Neurotrophin-3 contributes to benefits of human embryonic stem cell-derived cardiovascular progenitor cells against reperfused myocardial infarction. Stem Cells Transl Med. 2021;10:756–72.
Mersmann J, Zacharowski PA, Schmitz I, Zacharowski K. Caspase inhibitor zVAD.fmk reduces infarct size after myocardial ischaemia and reperfusion in rats but not in mice. Resuscitation. 2008;79:468–74.
Yaoita H, Ogawa K, Maehara K, Maruyama Y. Attenuation of ischemia/reperfusion injury in rats by a caspase inhibitor. Circulation. 1998;97:276–81.
Huang JQ, Radinovic S, Rezaiefar P, Black SC. In vivo myocardial infarct size reduction by a caspase inhibitor administered after the onset of ischemia. Eur J Pharmacol. 2000;402:139–42.
Souktani R, Pons S, Guegan C, Bouhidel O, Bruneval P, Zini R, et al. Cardioprotection against myocardial infarction with PTD-BIR3/RING, a XIAP mimicking protein. J Mol Cell Cardiol. 2009;46:713–8.
Inserte J, Cardona M, Poncelas-Nozal M, Hernando V, Vilardosa Ú, Aluja D, et al. Studies on the role of apoptosis after transient myocardial ischemia: genetic deletion of the executioner caspases-3 and -7 does not limit infarct size and ventricular remodeling. Basic Res Cardiol. 2016;111:18.
Weisleder N, Taffet GE, Capetanaki Y. Bcl-2 overexpression corrects mitochondrial defects and ameliorates inherited desmin null cardiomyopathy. Proc Natl Acad Sci USA. 2004;101:769–74.
Maloyan A, Sayegh J, Osinska H, Chua BH, Robbins J. Manipulation of death pathways in desmin-related cardiomyopathy. Circ Res. 2010;106:1524–32.
Khalil H, Peltzer N, Walicki J, Yang JY, Dubuis G, Gardiol N, et al. Caspase-3 protects stressed organs against cell death. Mol Cell Biol. 2012;32:4523–33.
Rodriguez-Ruiz ME, Buqué A, Hensler M, Chen J, Bloy N, Petroni G, et al. Apoptotic caspases inhibit abscopal responses to radiation and identify a new prognostic biomarker for breast cancer patients. Oncoimmunology. 2019;8:e1655964.
White MJ, McArthur K, Metcalf D, Lane RM, Cambier JC, Herold MJ, et al. Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell. 2014;159:1549–62.
Rongvaux A, Jackson R, Harman CC, Li T, West AP, de Zoete MR, et al. Apoptotic caspases prevent the induction of type I interferons by mitochondrial DNA. Cell. 2014;159:1563–77.
King KR, Aguirre AD, Ye YX, Sun Y, Roh JD, Ng RP Jr, et al. IRF3 and type I interferons fuel a fatal response to myocardial infarction. Nat Med. 2017;23:1481–7.
Fauvel H, Marchetti P, Chopin C, Formstecher P, Nevière R. Differential effects of caspase inhibitors on endotoxin-induced myocardial dysfunction and heart apoptosis. Am J Physiol Heart Circ Physiol. 2001;280:H1608–14.
Carlson DL, Maass DL, White J, Sikes P, Horton JW. Caspase inhibition reduces cardiac myocyte dyshomeostasis and improves cardiac contractile function after major burn injury. J Appl Physiol. 2007;103:323–30.
Araki T, Shibata M, Takano R, Hisahara S, Imamura S, Fukuuchi Y, et al. Conditional expression of anti-apoptotic protein p35 by Cre-mediated DNA recombination in cardiomyocytes from loxP-p35-transgenic mice. Cell Death Differ. 2000;7:485–92.
Amgalan D, Garner TP, Pekson R, Jia XF, Yanamandala M, Paulino V, et al. A small-molecule allosteric inhibitor of BAX protects against doxorubicin-induced cardiomyopathy. Nat Cancer. 2020;1:315–28.
Smyth LA, Meader L, Xiao F, Woodward M, Brady HJ, Lechler R, et al. Constitutive expression of the anti-apoptotic Bcl-2 family member A1 in murine endothelial cells leads to transplant tolerance. Clin Exp Immunol. 2017;188:219–25.
Grootaert MO, Schrijvers DM, Hermans M, Van Hoof VO, De Meyer GR, Martinet W. Caspase-3 deletion promotes necrosis in atherosclerotic plaques of ApoE knockout mice. Oxid Med Cell Longev. 2016;2016:3087469.
Chao ML, Guo J, Cheng WL, Zhu XY, She ZG, Huang Z, et al. Loss of caspase-activated DNase protects against atherosclerosis in apolipoprotein E-deficient mice. J Amn Heart Assoc. 2016;5:e004362.
Fontaine MAC, Westra MM, Bot I, Jin H, Franssen A, Bot M, et al. Low human and murine Mcl-1 expression leads to a pro-apoptotic plaque phenotype enriched in giant-cells. Sci Rep. 2019;9:14547.
Lee MKS, Kraakman MJ, Dragoljevic D, Hanssen NMJ, Flynn MC, Al-Sharea A, et al. Apoptotic ablation of platelets reduces atherosclerosis in mice with diabetes. Arterioscler Thromb Vasc Biol. 2021;41:1167–78.
Temmerman L, Westra MM, Bot I, van Vlijmen BJM, van Bree N, Bot M, et al. Leukocyte Bim deficiency does not impact atherogenesis in ldlr (−/−) mice, despite a pronounced induction of autoimmune inflammation. Sci Rep. 2017;7:3086.
Thorp E, Li Y, Bao L, Yao PM, Kuriakose G, Rong J, et al. Brief report: increased apoptosis in advanced atherosclerotic lesions of Apoe−/− mice lacking macrophage Bcl-2. Arterioscler Thromb Vasc Biol. 2009;29:169–72.
Wei Q, Dong G, Chen JK, Ramesh G. Dong Z. Bax and Bak have critical roles in ischemic acute kidney injury in global and proximal tubule-specific knockout mouse models. Kidney Int. 2013;84:138–48.
Wei Q, Yin XM, Wang MH, Dong Z. Bid deficiency ameliorates ischemic renal failure and delays animal death in C57BL/6 mice. Am J Physiol Ren Physiol. 2006;290:F35–42.
Chien CT, Shyue SK, Lai MK. Bcl-xL augmentation potentially reduces ischemia/reperfusion induced proximal and distal tubular apoptosis and autophagy. Transplantation. 2007;84:1183–90.
Mei S, Li L, Wei Q, Hao J, Su Y, Mei C, et al. Double knockout of Bax and Bak from kidney proximal tubules reduces unilateral urethral obstruction associated apoptosis and renal interstitial fibrosis. Sci Rep. 2017;7:44892.
Jang HS, Padanilam BJ. Simultaneous deletion of Bax and Bak is required to prevent apoptosis and interstitial fibrosis in obstructive nephropathy. Am J Physiol Ren Physiol. 2015;309:F540–550.
Yang B, Lan S, Dieudé M, Sabo-Vatasescu JP, Karakeussian-Rimbaud A, Turgeon J, et al. Caspase-3 is a pivotal regulator of microvascular rarefaction and renal fibrosis after ischemia-reperfusion injury. J Am Soc Nephrol. 2018;29:1900–16.
Lan S, Yang B, Migneault F, Turgeon J, Bourgault M, Dieudé M, et al. Caspase-3-dependent peritubular capillary dysfunction is pivotal for the transition from acute to chronic kidney disease after acute ischemia-reperfusion injury. Am J Physiol Ren Physiol. 2021;321:F335–f351.
Tao Y, Zafar I, Kim J, Schrier RW, Edelstein CL. Caspase-3 gene deletion prolongs survival in polycystic kidney disease. J Am Soc Nephrology. 2008;19:749–55.
Duplomb L, Droin N, Bouchot O, Thauvin-Robinet C, Bruel AL, Thevenon J, et al. A constitutive BCL2 down-regulation aggravates the phenotype of PKD1-mutant-induced polycystic kidney disease. Hum Mol Genet. 2017;26:4680–8.
Daemen MA, van ‘t Veer C, Denecker G, Heemskerk VH, Wolfs TG, Clauss M, et al. Inhibition of apoptosis induced by ischemia-reperfusion prevents inflammation. J Clin Investig. 1999;104:541–9.
Bral M, Pawlick R, Marfil-Garza B, Dadheech N, Hefler J, Thiesen A, et al. Pan-caspase inhibitor F573 mitigates liver ischemia reperfusion injury in a murine model. PLoS ONE. 2019;14:e0224567.
Tao Y, Kim J, Faubel S, Wu JC, Falk SA, Schrier RW, et al. Caspase inhibition reduces tubular apoptosis and proliferation and slows disease progression in polycystic kidney disease. Proc Natl Acad Sci USA. 2005;102:6954–9.
Yang B, Johnson TS, Haylor JL, Wagner B, Watson PF, El Kossi MM, et al. Effects of caspase inhibition on the progression of experimental glomerulonephritis. Kidney Int. 2003;63:2050–64.
Seery JP, Cattell V, Watt FM. Cutting edge: amelioration of kidney disease in a transgenic mouse model of lupus nephritis by administration of the caspase inhibitor carbobenzoxy-valyl-alanyl-aspartyl-(beta-o-methyl)-fluoromethylketone. J Immunol. 2001;167:2452–5.
Wen S, Wang ZH, Zhang CX, Yang Y, Fan QL. Caspase-3 promotes diabetic kidney disease through gasdermin E-mediated progression to secondary necrosis during apoptosis. Diabetes Metab Syndr Obes. 2020;13:313–23.
Belavgeni A, Meyer C, Stumpf J, Hugo C, Linkermann A. Ferroptosis and Necroptosis in the Kidney. Cell Chem Biol. 2020;27:448–62.
von Mässenhausen A, Tonnus W, Linkermann A. Cell death pathways drive necroinflammation during acute kidney injury. Nephron. 2018;140:144–7.
Guo R, Wang Y, Minto AW, Quigg RJ, Cunningham PN. Acute renal failure in endotoxemia is dependent on caspase activation. J Am Soc Nephrology. 2004;15:3093–102.
Herzog C, Yang C, Holmes A, Kaushal GP. zVAD-fmk prevents cisplatin-induced cleavage of autophagy proteins but impairs autophagic flux and worsens renal function. Am J Physiol Ren Physiol. 2012;303:F1239–50.
Linkermann A, Heller JO, Prókai A, Weinberg JM, De Zen F, Himmerkus N, et al. The RIP1-kinase inhibitor necrostatin-1 prevents osmotic nephrosis and contrast-induced AKI in mice. J Am Soc Nephrology. 2013;24:1545–57.
Brinkmann K, Waring P, Glaser SP, Wimmer V, Cottle DL, Tham MS, et al. BCL-XL exerts a protective role against anemia caused by radiation-induced kidney damage. EMBO J. 2020;39:e105561.
Yin XM, Wang K, Gross A, Zhao Y, Zinkel S, Klocke B, et al. Bid-deficient mice are resistant to Fas-induced hepatocellular apoptosis. Nature. 1999;400:886–91.
DuBray BJ Jr, Conzen KD, Upadhya GA, Gunter KL, Jia J, Knolhoff BL, et al. BH3-only proteins contribute to steatotic liver ischemia-reperfusion injury. J Surg Res. 2015;194:653–8.
Selzner M, Rüdiger HA, Selzner N, Thomas DW, Sindram D, Clavien PA. Transgenic mice overexpressing human Bcl-2 are resistant to hepatic ischemia and reperfusion. J Hepatol. 2002;36:218–25.
Cursio R, Gugenheim J, Ricci JE, Crenesse D, Rostagno P, Maulon L, et al. Caspase inhibition protects from liver injury following ischemia and reperfusion in rats. Transpl Int. 2000;13:S568–72.
Kaufmann T, Jost PJ, Pellegrini M, Puthalakath H, Gugasyan R, Gerondakis S, et al. Fatal hepatitis mediated by tumor necrosis factor TNFalpha requires caspase-8 and involves the BH3-only proteins Bid and Bim. Immunity. 2009;30:56–66.
Riddle-Taylor E, Nagasaki K, Lopez J, Esquivel CO, Martinez OM, Krams SM. Mutations to bid cleavage sites protect hepatocytes from apoptosis after ischemia/reperfusion injury. Transplantation. 2007;84:778–85.
Lauer C, Brunner T, Corazza N. The proapoptotic Bcl-2 family member Bim plays a central role during the development of virus-induced hepatitis. J Immunol. 2012;188:916–22.
Chen D, Ni HM, Wang L, Ma X, Yu J, Ding WX, et al. p53 Up-regulated modulator of apoptosis induction mediates acetaminophen-induced necrosis and liver injury in mice. Hepatology. 2019;69:2164–79.
Badmann A, Keough A, Kaufmann T, Bouillet P, Brunner T, Corazza N. Role of TRAIL and the pro-apoptotic Bcl-2 homolog Bim in acetaminophen-induced liver damage. Cell Death Dis. 2011;2:e171.
Naim S, Fernandez-Marrero Y, de Brot S, Bachmann D, Kaufmann T. Loss of BOK has a minor impact on acetaminophen overdose-induced liver damage in mice. Int J Mol Sci. 2021;22:3281.
Yoshida N, Iwata H, Yamada T, Sekino T, Matsuo H, Shirahashi K, et al. Improvement of the survival rate after rat massive hepatectomy due to the reduction of apoptosis by caspase inhibitor. J Gastroenterol Hepatol. 2007;22:2015–21.
Roychowdhury S, Chiang DJ, Mandal P, McMullen MR, Liu X, Cohen JI, et al. Inhibition of apoptosis protects mice from ethanol-mediated acceleration of early markers of CCl4 -induced fibrosis but not steatosis or inflammation. Alcohol, Clin Exp Res. 2012;36:1139–47.
Eguchi A, De Mollerat Du Jeu X, Johnson CD, Nektaria A, Feldstein AE. Liver Bid suppression for treatment of fibrosis associated with non-alcoholic steatohepatitis. J Hepatol. 2016;64:699–707.
Higuchi H, Miyoshi H, Bronk SF, Zhang H, Dean N, Gores GJ. Bid antisense attenuates bile acid-induced apoptosis and cholestatic liver injury. J Pharmacol Exp Ther. 2001;299:866–73.
Nalapareddy P, Schüngel S, Hong JY, Manns MP, Jaeschke H, Vogel A. The BH3-only protein bid does not mediate death-receptor-induced liver injury in obstructive cholestasis. Am J Pathol. 2009;175:1077–85.
Kahraman A, Mott JL, Bronk SF, Werneburg NW, Barreyro FJ, Guicciardi ME, et al. Overexpression of mcl-1 attenuates liver injury and fibrosis in the bile duct-ligated mouse. Digest Dis Sci. 2009;54:1908–17.
Mitchell C, Mahrouf-Yorgov M, Mayeuf A, Robin MA, Mansouri A, Fromenty B, et al. Overexpression of Bcl-2 in hepatocytes protects against injury but does not attenuate fibrosis in a mouse model of chronic cholestatic liver disease. Lab Investig. 2011;91:273–82.
He L, Sehrawat TS, Verma VK, Navarro-Corcuera A, Sidhu G, Mauer A, et al. XIAP knockdown in alcohol-associated liver disease models exhibits divergent in vitro and in vivo phenotypes owing to a potential zonal inhibitory role of SMAC. Front Physiol. 2021;12:664222.
Zilu S, Qian H, Haibin W, Chenxu G, Deshuai L, Qiang L, et al. Effects of XIAP on high fat diet-induced hepatic steatosis: a mechanism involving NLRP3 inflammasome and oxidative stress. Aging. 2019;11:12177–201.
Thapaliya S, Wree A, Povero D, Inzaugarat ME, Berk M, Dixon L, et al. Caspase 3 inactivation protects against hepatic cell death and ameliorates fibrogenesis in a diet-induced NASH model. Digest Dis Sci. 2014;59:1197–206.
Weng SY, Yang CY, Li CC, Sun TP, Tung SY, Yen JJ, et al. Synergism between p53 and Mcl-1 in protecting from hepatic injury, fibrosis and cancer. J Hepatol. 2011;54:685–94.
Hikita H, Kodama T, Shimizu S, Li W, Shigekawa M, Tanaka S, et al. Bak deficiency inhibits liver carcinogenesis: a causal link between apoptosis and carcinogenesis. J Hepatol. 2012;57:92–100.
Rabachini T, Fernandez-Marrero Y, Montani M, Loforese G, Sladky V, He Z, et al. BOK promotes chemical-induced hepatocarcinogenesis in mice. Cell Death Differ. 2018;25:708–20.
Wree A, Johnson CD, Font-Burgada J, Eguchi A, Povero D, Karin M, et al. Hepatocyte-specific Bid depletion reduces tumor development by suppressing inflammation-related compensatory proliferation. Cell Death Differ. 2015;22:1985–94.
Orlik J, Schüngel S, Buitrago-Molina LE, Marhenke S, Geffers R, Endig J, et al. The BH3-only protein BID impairs the p38-mediated stress response and promotes hepatocarcinogenesis during chronic liver injury in mice. Hepatology. 2015;62:816–28.
Kim JY, Garcia-Carbonell R, Yamachika S, Zhao P, Dhar D, Loomba R, et al. ER stress drives lipogenesis and steatohepatitis via caspase-2 activation of S1P. Cell. 2018;175:133–145.e115.
Machado MV, Michelotti GA, Jewell ML, Pereira TA, Xie G, Premont RT, et al. Caspase-2 promotes obesity, the metabolic syndrome and nonalcoholic fatty liver disease. Cell Death Dis. 2016;7:e2096.
Barreyro FJ, Holod S, Finocchietto PV, Camino AM, Aquino JB, Avagnina A, et al. The pan-caspase inhibitor Emricasan (IDN-6556) decreases liver injury and fibrosis in a murine model of non-alcoholic steatohepatitis. Liver Int. 2015;35:953–66.
Witek RP, Stone WC, Karaca FG, Syn WK, Pereira TA, Agboola KM, et al. Pan-caspase inhibitor VX-166 reduces fibrosis in an animal model of nonalcoholic steatohepatitis. Hepatology. 2009;50:1421–30.
Gracia-Sancho J, Manicardi N, Ortega-Ribera M, Maeso-Díaz R, Guixé-Muntet S, Fernández-Iglesias A, et al. Emricasan ameliorates portal hypertension and liver fibrosis in cirrhotic rats through a hepatocyte-mediated paracrine mechanism. Hepatol Commun. 2019;3:987–1000.
Eguchi A, Koyama Y, Wree A, Johnson CD, Nakamura R, Povero D, et al. Emricasan, a pan-caspase inhibitor, improves survival and portal hypertension in a murine model of common bile-duct ligation. J Mol Med. 2018;96:575–83.
Canbay A, Feldstein A, Baskin-Bey E, Bronk SF, Gores GJ. The caspase inhibitor IDN-6556 attenuates hepatic injury and fibrosis in the bile duct ligated mouse. J Pharmacol Exp Ther. 2004;308:1191–6.
Garcia-Tsao G, Fuchs M, Shiffman M, Borg BB, Pyrsopoulos N, Shetty K, et al. Emricasan (IDN-6556) Lowers Portal Pressure in Patients With Compensated Cirrhosis and Severe Portal Hypertension. Hepatology. 2019;69:717–28.
Frenette C, Kayali Z, Mena E, Mantry PS, Lucas KJ, Neff G, et al. Emricasan to prevent new decompensation in patients with NASH-related decompensated cirrhosis. J Hepatol. 2021;74:274–82.
Xu WF, Zhang Q, Ding CJ, Sun HY, Che Y, Huang H, et al. Gasdermin E-derived caspase-3 inhibitors effectively protect mice from acute hepatic failure. Acta Pharmacol Sin. 2021;42:68–76.
Högstrand K, Hejll E, Sander B, Rozell B, Larsson LG, Grandien A. Inhibition of the intrinsic but not the extrinsic apoptosis pathway accelerates and drives MYC-driven tumorigenesis towards acute myeloid leukemia. PLoS ONE. 2012;7:e31366.
Finch A, Prescott J, Shchors K, Hunt A, Soucek L, Dansen TB, et al. Bcl-xL gain of function and p19 ARF loss of function cooperate oncogenically with Myc in vivo by distinct mechanisms. Cancer Cell. 2006;10:113–20.
Swanson PJ, Kuslak SL, Fang W, Tze L, Gaffney P, Selby S, et al. Fatal acute lymphoblastic leukemia in mice transgenic for B cell-restricted bcl-xL and c-myc. J Immunol. 2004;172:6684–91.
Strasser A, Harris AW, Bath ML, Cory S. Novel primitive lymphoid tumours induced in transgenic mice by cooperation between myc and bcl-2. Nature. 1990;348:331–3.
Campbell KJ, Bath ML, Turner ML, Vandenberg CJ, Bouillet P, Metcalf D, et al. Elevated Mcl-1 perturbs lymphopoiesis, promotes transformation of hematopoietic stem/progenitor cells, and enhances drug resistance. Blood. 2010;116:3197–207.
Kelly GL, Grabow S, Glaser SP, Fitzsimmons L, Aubrey BJ, Okamoto T, et al. Targeting of MCL-1 kills MYC-driven mouse and human lymphomas even when they bear mutations in p53. Genes Dev. 2014;28:58–70.
Vandenberg CJ, Cory S. ABT-199, a new Bcl-2-specific BH3 mimetic, has in vivo efficacy against aggressive Myc-driven mouse lymphomas without provoking thrombocytopenia. Blood. 2013;121:2285–8.
Kelly PN, Grabow S, Delbridge AR, Adams JM, Strasser A. Prophylactic treatment with the BH3 mimetic ABT-737 impedes Myc-driven lymphomagenesis in mice. Cell Death Differ. 2013;20:57–63.
Mason KD, Vandenberg CJ, Scott CL, Wei AH, Cory S, Huang DC, et al. In vivo efficacy of the Bcl-2 antagonist ABT-737 against aggressive Myc-driven lymphomas. Proc Natl Acad Sci USA. 2008;105:17961–6.
Yin K, Lee J, Liu Z, Kim H, Martin DR, Wu D, et al. Mitophagy protein PINK1 suppresses colon tumor growth by metabolic reprogramming via p53 activation and reducing acetyl-CoA production. Cell Death Differ. 2021;28:2421–35.
Bowen ME, Mulligan AS, Sorayya A, Attardi LD. Puma- and Caspase9-mediated apoptosis is dispensable for p53-driven neural crest-based developmental defects. Cell Death Differ. 2021;28:2083–94.
Liang J, Niu Z, Zhang B, Yu X, Zheng Y, Wang C, et al. p53-dependent elimination of aneuploid mitotic offspring by entosis. Cell Death Differ. 2021;28:799–813.
Fischer M, Steiner L, Engeland K. The transcription factor p53: not a repressor, solely an activator. Cell Cycle. 2014;13:3037–58.
Engeland K. Cell cycle arrest through indirect transcriptional repression by p53: I have a DREAM. Cell Death Differ. 2018;25:114–32.
Engeland K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022;29:946–60.
Aubrey BJ, Kelly GL, Janic A, Herold MJ, Strasser A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018;25:104–13.
Vogelstein B, Lane D, Levine AJ. Surfing the p53 network. Nature. 2000;408:307–10.
Uxa S, Castillo-Binder P, Kohler R, Stangner K, Muller GA, Engeland K. Ki-67 gene expression. Cell Death Differ. 2021;28:3357–70.
Kelly PN, Grabow S, Delbridge AR, Strasser A, Adams JM. Endogenous Bcl-xL is essential for Myc-driven lymphomagenesis in mice. Blood. 2011;118:6380–6.
Kelly PN, Puthalakath H, Adams JM, Strasser A. Endogenous bcl-2 is not required for the development of Emu-myc-induced B-cell lymphoma. Blood. 2007;109:4907–13.
Grabow S, Delbridge AR, Aubrey BJ, Vandenberg CJ, Strasser A. Loss of a single Mcl-1 allele inhibits MYC-driven lymphomagenesis by sensitizing Pro-B cells to apoptosis. Cell Rep. 2016;14:2337–47.
Xiang Z, Luo H, Payton JE, Cain J, Ley TJ, Opferman JT, et al. Mcl1 haploinsufficiency protects mice from Myc-induced acute myeloid leukemia. J Clin Investig. 2010;120:2109–18.
Grabow S, Kelly GL, Delbridge AR, Kelly PN, Bouillet P, Adams JM, et al. Critical B-lymphoid cell intrinsic role of endogenous MCL-1 in c-MYC-induced lymphomagenesis. Cell Death Dis. 2016;7:e2132.
Grabow S, Delbridge AR, Valente LJ, Strasser A. MCL-1 but not BCL-XL is critical for the development and sustained expansion of thymic lymphoma in p53-deficient mice. Blood. 2014;124:3939–46.
Grabow S, Waring P, Happo L, Cook M, Mason KD, Kelly PN, et al. Pharmacological blockade of Bcl-2, Bcl-x(L) and Bcl-w by the BH3 mimetic ABT-737 has only minor impact on tumour development in p53-deficient mice. Cell Death Differ. 2012;19:623–32.
Beverly LJ, Varmus HE. MYC-induced myeloid leukemogenesis is accelerated by all six members of the antiapoptotic BCL family. Oncogene. 2009;28:1274–9.
Diepstraten ST, Chang C, Tai L, Gong JN, Lan P, Dowell AC, et al. BCL-W is dispensable for the sustained survival of select Burkitt lymphoma and diffuse large B-cell lymphoma cell lines. Blood Adv. 2020;4:356–66.
Glaser SP, Lee EF, Trounson E, Bouillet P, Wei A, Fairlie WD, et al. Anti-apoptotic Mcl-1 is essential for the development and sustained growth of acute myeloid leukemia. Genes Dev. 2012;26:120–5.
Kotschy A, Szlavik Z, Murray J, Davidson J, Maragno AL, Le Toumelin-Braizat G, et al. The MCL1 inhibitor S63845 is tolerable and effective in diverse cancer models. Nature. 2016;538:477–82.
Adams CM, Kim AS, Mitra R, Choi JK, Gong JZ, Eischen CM. BCL-W has a fundamental role in B cell survival and lymphomagenesis. J Clin Investig. 2017;127:635–50.
Eischen CM, Roussel MF, Korsmeyer SJ, Cleveland JL. Bax loss impairs Myc-induced apoptosis and circumvents the selection of p53 mutations during Myc-mediated lymphomagenesis. Mol Cell Biol. 2001;21:7653–62.
Egle A, Harris AW, Bouillet P, Cory S. Bim is a suppressor of Myc-induced mouse B cell leukemia. Proc Natl Acad Sci USA. 2004;101:6164–9.
Delbridge AR, Grabow S, Bouillet P, Adams JM, Strasser A. Functional antagonism between pro-apoptotic BIM and anti-apoptotic BCL-XL in MYC-induced lymphomagenesis. Oncogene. 2015;34:1872–6.
Frenzel A, Labi V, Chmelewskij W, Ploner C, Geley S, Fiegl H, et al. Suppression of B-cell lymphomagenesis by the BH3-only proteins Bmf and Bad. Blood. 2010;115:995–1005.
Hemann MT, Zilfou JT, Zhao Z, Burgess DJ, Hannon GJ, Lowe SW. Suppression of tumorigenesis by the p53 target PUMA. Proc Natl Acad Sci USA. 2004;101:9333–8.
Michalak EM, Jansen ES, Happo L, Cragg MS, Tai L, Smyth GK, et al. Puma and to a lesser extent Noxa are suppressors of Myc-induced lymphomagenesis. Cell Death Differ. 2009;16:684–96.
Garrison SP, Jeffers JR, Yang C, Nilsson JA, Hall MA, Rehg JE, et al. Selection against PUMA gene expression in Myc-driven B-cell lymphomagenesis. Mol Cell Biol. 2008;28:5391–402.
Mérino D, Strasser A, Bouillet P. Bim must be able to engage all pro-survival Bcl-2 family members for efficient tumor suppression. Oncogene. 2012;31:3392–6.
Shang Q, Zhang D, Guo C, Lin Q, Guo Z, Deng C. Potential synergism of Bim with p53 in mice with Myc‑induced lymphoma in a mouse lymphoma model. Mol Med Rep. 2012;5:1401–8.
Delbridge AR, Pang SH, Vandenberg CJ, Grabow S, Aubrey BJ, Tai L, et al. RAG-induced DNA lesions activate proapoptotic BIM to suppress lymphomagenesis in p53-deficient mice. J Exp Med. 2016;213:2039–48.
Knudson CM, Johnson GM, Lin Y, Korsmeyer SJ. Bax accelerates tumorigenesis in p53-deficient mice. Cancer Res. 2001;61:659–65.
Valente LJ, Grabow S, Vandenberg CJ, Strasser A, Janic A. Combined loss of PUMA and p21 accelerates c-MYC-driven lymphoma development considerably less than loss of one allele of p53. Oncogene. 2016;35:3866–71.
Happo L, Phipson B, Smyth GK, Strasser A, Scott CL. Neither loss of Bik alone, nor combined loss of Bik and Noxa, accelerate murine lymphoma development or render lymphoma cells resistant to DNA damaging drugs. Cell Death Dis. 2012;3:e306.
Ho LH, Taylor R, Dorstyn L, Cakouros D, Bouillet P, Kumar S. A tumor suppressor function for caspase-2. Proc Natl Acad Sci USA. 2009;106:5336–41.
Scott CL, Schuler M, Marsden VS, Egle A, Pellegrini M, Nesic D, et al. Apaf-1 and caspase-9 do not act as tumor suppressors in myc-induced lymphomagenesis or mouse embryo fibroblast transformation. J Cell Biol. 2004;164:89–96.
Guirguis AA, Slape CI, Failla LM, Saw J, Tremblay CS, Powell DR, et al. PUMA promotes apoptosis of hematopoietic progenitors driving leukemic progression in a mouse model of myelodysplasia. Cell Death Differ. 2016;23:1049–59.
Michalak EM, Vandenberg CJ, Delbridge AR, Wu L, Scott CL, Adams JM, et al. Apoptosis-promoted tumorigenesis: gamma-irradiation-induced thymic lymphomagenesis requires Puma-driven leukocyte death. Genes Dev. 2010;24:1608–13.
Labi V, Erlacher M, Krumschnabel G, Manzl C, Tzankov A, Pinon J, et al. Apoptosis of leukocytes triggered by acute DNA damage promotes lymphoma formation. Genes Dev. 2010;24:1602–7.
Slinger E, Wensveen FM, Guikema JE, Kater AP, Eldering E. Chronic lymphocytic leukemia development is accelerated in mice with deficiency of the pro-apoptotic regulator NOXA. Haematologica. 2016;101:e374–77.
Katz SG, Labelle JL, Meng H, Valeriano RP, Fisher JK, Sun H, et al. Mantle cell lymphoma in cyclin D1 transgenic mice with Bim-deficient B cells. Blood. 2014;123:884–93.
Anstee NS, Bilardi RA, Ng AP, Xu Z, Robati M, Vandenberg CJ, et al. Impact of elevated anti-apoptotic MCL-1 and BCL-2 on the development and treatment of MLL-AF9 AML in mice. Cell Death Differ. 2019;26:1316–31.
Vandenberg CJ, Waring P, Strasser A, Cory S. Plasmacytomagenesis in Eμ-v-abl transgenic mice is accelerated when apoptosis is restrained. Blood. 2014;124:1099–109.
Spinner S, Crispatzu G, Yi JH, Munkhbaatar E, Mayer P, Höckendorf U, et al. Re-activation of mitochondrial apoptosis inhibits T-cell lymphoma survival and treatment resistance. Leukemia. 2016;30:1520–30.
Puccini J, Shalini S, Voss AK, Gatei M, Wilson CH, Hiwase DK, et al. Loss of caspase-2 augments lymphomagenesis and enhances genomic instability in Atm-deficient mice. Proc Natl Acad Sci USA. 2013;110:19920–5.
Fava LL, Schuler F, Sladky V, Haschka MD, Soratroi C, Eiterer L, et al. The PIDDosome activates p53 in response to supernumerary centrosomes. Genes Dev. 2017;31:34–45.
Lachowiez C, DiNardo CD, Konopleva M. Venetoclax in acute myeloid leukemia - current and future directions. Leuk lymphoma. 2020;61:1313–22.
Nechiporuk T, Kurtz SE, Nikolova O, Liu T, Jones CL, D’Alessandro A, et al. The TP53 apoptotic network is a primary mediator of resistance to BCL2 inhibition in AML cells. Cancer Discov. 2019;9:910–25.
Bosc C, Saland E, Bousard A, Gadaud N, Sabatier M, Cognet G, et al. Mitochondrial inhibitors circumvent adaptive resistance to venetoclax and cytarabine combination therapy in acute myeloid leukemia. Nat Cancer. 2021;2:1204–23.
Thijssen R, Diepstraten ST, Moujalled D, Chew E, Flensburg C, Shi MX, et al. Intact TP-53 function is essential for sustaining durable responses to BH3-mimetic drugs in leukemias. Blood. 2021;137:2721–35.
Thomalla D, Beckmann L, Grimm C, Oliverio M, Meder L, Herling CD, et al. Deregulation and epigenetic modification of BCL2-family genes cause resistance to venetoclax in hematologic malignancies. Blood. 2022;140:2113–26.
Jilg S, Reidel V, Müller-Thomas C, König J, Schauwecker J, Höckendorf U, et al. Blockade of BCL-2 proteins efficiently induces apoptosis in progenitor cells of high-risk myelodysplastic syndromes patients. Leukemia. 2016;30:112–23.
Ganan-Gomez I, Yang H, Ma F, Montalban-Bravo G, Thongon N, Marchica V, et al. Stem cell architecture drives myelodysplastic syndrome progression and predicts response to venetoclax-based therapy. Nat Med. 2022;28:557–67.
Jilg S, Hauch RT, Kauschinger J, Buschhorn L, Odinius TO, Dill V, et al. Venetoclax with azacitidine targets refractory MDS but spares healthy hematopoiesis at tailored dose. Exp Hematol Oncol. 2019;8:9.
Jager R, Herzer U, Schenkel J, Weiher H. Overexpression of Bcl-2 inhibits alveolar cell apoptosis during involution and accelerates c-myc-induced tumorigenesis of the mammary gland in transgenic mice. Oncogene. 1997;15:1787–95.
Shibata MA, Liu ML, Knudson MC, Shibata E, Yoshidome K, Bandey T, et al. Haploid loss of bax leads to accelerated mammary tumor development in C3(1)/SV40-TAg transgenic mice: reduction in protective apoptotic response at the preneoplastic stage. EMBO J. 1999;18:2692–701.
Jamerson MH, Johnson MD, Korsmeyer SJ, Furth PA, Dickson RB. Bax regulates c-Myc-induced mammary tumour apoptosis but not proliferation in MMTV-c-myc transgenic mice. Br J Cancer. 2004;91:1372–9.
Bean GR, Ganesan YT, Dong Y, Takeda S, Liu H, Chan PM, et al. PUMA and BIM are required for oncogene inactivation-induced apoptosis. Sci Signal. 2013;6:ra20.
Parsons MJ, McCormick L, Janke L, Howard A, Bouchier-Hayes L, Green DR. Genetic deletion of caspase-2 accelerates MMTV/c-neu-driven mammary carcinogenesis in mice. Cell Death Differ. 2013;20:1174–82.
Murphy KL, Kittrell FS, Gay JP, Jäger R, Medina D, Rosen JM. Bcl-2 expression delays mammary tumor development in dimethylbenz(a)anthracene-treated transgenic mice. Oncogene. 1999;18:6597–604.
van der Heijden M, Zimberlin CD, Nicholson AM, Colak S, Kemp R, Meijer SL, et al. Bcl-2 is a critical mediator of intestinal transformation. Nat Commun. 2016;7:10916.
Scherr AL, Gdynia G, Salou M, Radhakrishnan P, Duglova K, Heller A, et al. Bcl-xL is an oncogenic driver in colorectal cancer. Cell Death Dis. 2016;7:e2342.
Qiu W, Carson-Walter EB, Kuan SF, Zhang L, Yu J. PUMA suppresses intestinal tumorigenesis in mice. Cancer Res. 2009;69:4999–5006.
Muthalagu N, Junttila MR, Wiese KE, Wolf E, Morton J, Bauer B, et al. BIM is the primary mediator of MYC-induced apoptosis in multiple solid tissues. Cell Rep. 2014;8:1347–53.
Ramesh P, Lannagan TRM, Jackstadt R, Atencia Taboada L, Lansu N, Wirapati P, et al. BCL-XL is crucial for progression through the adenoma-to-carcinoma sequence of colorectal cancer. Cell Death Differ. 2021;28:3282–96.
Zeuner A, Francescangeli F, Contavalli P, Zapparelli G, Apuzzo T, Eramo A, et al. Elimination of quiescent/slow-proliferating cancer stem cells by Bcl-XL inhibition in non-small cell lung cancer. Cell Death Differ. 2014;21:1877–88.
Colak S, Zimberlin CD, Fessler E, Hogdal L, Prasetyanti PR, Grandela CM, et al. Decreased mitochondrial priming determines chemoresistance of colon cancer stem cells. Cell Death Differ. 2014;21:1170–7.
Garcia I, Crowther AJ, Gama V, Miller CR, Deshmukh M, Gershon TR. Bax deficiency prolongs cerebellar neurogenesis, accelerates medulloblastoma formation and paradoxically increases both malignancy and differentiation. Oncogene. 2013;32:2304–14.
Yin C, Knudson CM, Korsmeyer SJ, Van Dyke T. Bax suppresses tumorigenesis and stimulates apoptosis in vivo. Nature. 1997;385:637–40.
Terry MR, Arya R, Mukhopadhyay A, Berrett KC, Clair PM, Witt B, et al. Caspase-2 impacts lung tumorigenesis and chemotherapy response in vivo. Cell Death Differ. 2015;22:719–30.
Munkhbaatar E, Dietzen M, Agrawal D, Anton M, Jesinghaus M, Boxberg M, et al. MCL-1 gains occur with high frequency in lung adenocarcinoma and can be targeted therapeutically. Nat Commun. 2020;11:4527.
Meinhardt AL, Munkhbaatar E, Höckendorf U, Dietzen M, Dechant M, Anton M, et al. The BCL-2 family member BOK promotes KRAS-driven lung cancer progression in a p53-dependent manner. Oncogene. 2022;41:1376–82.
He M, Chaurushiya MS, Webster JD, Kummerfeld S, Reja R, Chaudhuri S, et al. Intrinsic apoptosis shapes the tumor spectrum linked to inactivation of the deubiquitinase BAP1. Sciene. 2019;364:283–5.
Carbone M, Harbour JW, Brugarolas J, Bononi A, Pagano I, Dey A, et al. Biological mechanisms and clinical significance of BAP1 mutations in human cancer. Cancer Discov. 2020;10:1103–20.
Novelli F, Bononi A, Wang Q, Bai F, Patergnani S, Kricek F, et al. BAP1 forms a trimer with HMGB1 and HDAC1 that modulates gene × environment interaction with asbestos. Proc Natl Acad Sci USA. 2021;118:e2111946118.
Bononi A, Giorgi C, Patergnani S, Larson D, Verbruggen K, Tanji M, et al. BAP1 regulates IP3R3-mediated Ca(2+) flux to mitochondria suppressing cell transformation. Nature. 2017;546:549–53.
Dansen TB, Whitfield J, Rostker F, Brown-Swigart L, Evan GI. Specific requirement for Bax, not Bak, in Myc-induced apoptosis and tumor suppression in vivo. J Biol Chem. 2006;281:10890–5.
Radziszewska A, Schroer SA, Choi D, Tajmir P, Radulovich N, Ho JC, et al. Absence of caspase-3 protects pancreatic {beta}-cells from c-Myc-induced apoptosis without leading to tumor formation. J Biol Chem. 2009;284:10947–56.
Evan GI, Christophorou M, Lawlor EA, Ringshausen I, Prescott J, Dansen T, et al. Oncogene-dependent tumor suppression: using the dark side of the force for cancer therapy. Cold Spring Harb Symp Quant Biol. 2005;70:263–73.
Shalini S, Nikolic A, Wilson CH, Puccini J, Sladojevic N, Finnie J, et al. Caspase-2 deficiency accelerates chemically induced liver cancer in mice. Cell Death Differ. 2016;23:1727–36.
Qiu W, Wang X, Leibowitz B, Yang W, Zhang L, Yu J. PUMA-mediated apoptosis drives chemical hepatocarcinogenesis in mice. Hepatology. 2011;54:1249–58.
Pierce RH, Vail ME, Ralph L, Campbell JS, Fausto N. Bcl-2 expression inhibits liver carcinogenesis and delays the development of proliferating foci. Am J Pathol. 2002;160:1555–60.
Vail ME, Pierce RH, Fausto N. Bcl-2 delays and alters hepatic carcinogenesis induced by transforming growth factor alpha. Cancer Res. 2001;61:594–601.
Pena JC, Rudin CM, Thompson CB. A Bcl-xL transgene promotes malignant conversion of chemically initiated skin papillomas. Cancer Res. 1998;58:2111–6.
Schenkel J, Weiher H, Fürstenberger G, Jäger R. Suprabasal BCL-2 expression does not sensitize to chemically-induced skin cancer in transgenic mice. Anticancer Res. 2008;28:2825–9.
Rossiter H, Beissert S, Mayer C, Schön MP, Wienrich BG, Tschachler E, et al. Targeted expression of bcl-2 to murine basal epidermal keratinocytes results in paradoxical retardation of ultraviolet- and chemical-induced tumorigenesis. Cancer Res. 2001;61:3619–26.
Kim DJ, Kataoka K, Sano S, Connolly K, Kiguchi K, DiGiovanni J. Targeted disruption of Bcl-xL in mouse keratinocytes inhibits both UVB- and chemically induced skin carcinogenesis. Mol Carcinogen. 2009;48:873–85.
Strasser A, Whittingham S, Vaux DL, Bath ML, Adams JM, Cory S, et al. Enforced BCL2 expression in B-lymphoid cells prolongs antibody responses and elicits autoimmune disease. Proc Natl Acad Sci USA. 1991;88:8661–5.
Bouillet P, Metcalf D, Huang DC, Tarlinton DM, Kay TW, Köntgen F, et al. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science. 1999;286:1735–8.
Mason KD, Lin A, Robb L, Josefsson EC, Henley KJ, Gray DH, et al. Proapoptotic Bak and Bax guard against fatal systemic and organ-specific autoimmune disease. Proc Natl Acad Sci USA. 2013;110:2599–604.
Scatizzi JC, Bickel E, Hutcheson J, Haines GK 3rd, Perlman H. Bim deficiency leads to exacerbation and prolongation of joint inflammation in experimental arthritis. Arthritis Rheum. 2006;54:3182–93.
Li J, Zhang L, Zheng Y, Shao R, Liang Q, Yu W, et al. BAD inactivation exacerbates rheumatoid arthritis pathology by promoting survival of sublining macrophages. eLife. 2020;9:e56309.
Scatizzi JC, Hutcheson J, Bickel E, Haines GK 3rd, Perlman H. Pro-apoptotic Bid is required for the resolution of the effector phase of inflammatory arthritis. Arthritis Res Ther. 2007;9:R49.
Scatizzi JC, Hutcheson J, Pope RM, Firestein GS, Koch AE, Mavers M, et al. Bim-Bcl-2 homology 3 mimetic therapy is effective at suppressing inflammatory arthritis through the activation of myeloid cell apoptosis. Arthritis Rheum. 2010;62:441–51.
Moore CS, Hebb AL, Blanchard MM, Crocker CE, Liston P, Korneluk RG, et al. Increased X-linked inhibitor of apoptosis protein (XIAP) expression exacerbates experimental autoimmune encephalomyelitis (EAE). J Neuroimmunol. 2008;203:79–93.
Lev N, Barhum Y, Melamed E, Offen D. Bax-ablation attenuates experimental autoimmune encephalomyelitis in mice. Neurosci Lett. 2004;359:139–42.
Ludwinski MW, Sun J, Hilliard B, Gong S, Xue F, Carmody RJ, et al. Critical roles of Bim in T cell activation and T cell-mediated autoimmune inflammation in mice. J Clin Investig. 2009;119:1706–13.
Offen D, Kaye JF, Bernard O, Merims D, Coire CI, Panet H, et al. Mice overexpressing Bcl-2 in their neurons are resistant to myelin oligodendrocyte glycoprotein (MOG)-induced experimental autoimmune encephalomyelitis (EAE). J Mol Neurosci. 2000;15:167–76.
Lagares D, Santos A, Grasberger PE, Liu F, Probst CK, Rahimi RA, et al. Targeted apoptosis of myofibroblasts with the BH3 mimetic ABT-263 reverses established fibrosis. Sci Transl Med. 2017;9:eaal3765.
Sun J, Mao LQ, Polonsky KS, Ren DC. Pancreatic β-cell death due to Pdx-1 deficiency requires multi-BH domain protein Bax but Not Bak. J Biol Chem. 2016;291:13529–34.
White SA, Zhang LS, Pasula DJ, Yang YHC, Luciani DS. Bax and Bak jointly control survival and dampen the early unfolded protein response in pancreatic β-cells under glucolipotoxic stress. Sci Rep. 2020;10:10986.
Krishnamurthy B, Chee J, Jhala G, Trivedi P, Catterall T, Selck C, et al. BIM deficiency protects NOD mice from diabetes by diverting thymocytes to regulatory T cells. Diabetes. 2015;64:3229–38.
Ren D, Sun J, Wang C, Ye H, Mao L, Cheng EH, et al. Role of BH3-only molecules Bim and Puma in β-cell death in Pdx1 deficiency. Diabetes. 2014;63:2744–50.
Ren D, Sun J, Mao L, Ye H, Polonsky KS. BH3-only molecule Bim mediates β-cell death in IRS2 deficiency. Diabetes. 2014;63:3378–87.
Pfeiffer S, Halang L, Düssmann H, Byrne MM, Prehn J. BH3-Only protein bmf is required for the maintenance of glucose homeostasis in an in vivo model of HNF1α-MODY diabetes. Cell Death Discov. 2015;1:15041.
Uhlemeyer C, Muller N, Rieck M, Kuboth J, Schlegel C, Griess K, et al. Selective ablation of P53 in pancreatic beta cells fails to ameliorate glucose metabolism in genetic, dietary and pharmacological models of diabetes mellitus. Mol Metab. 2023;67:101650.
Rohner L, Reinhart R, Hagmann B, Odermatt A, Babirye A, Kaufmann T, et al. FcɛRI cross-linking and IL-3 protect human basophils from intrinsic apoptotic stress. J Allergy Clin Immunol. 2018;142:1647–1650.e1643.
Reinhart R, Kaufmann T. IL-4 enhances survival of in vitro-differentiated mouse basophils through transcription-independent signaling downstream of PI3K. Cell Death Dis. 2018;9:713.
Didichenko SA, Spiegl N, Brunner T, Dahinden CA. IL-3 induces a Pim1-dependent antiapoptotic pathway in primary human basophils. Blood. 2008;112:3949–58.
Vassina EM, Yousefi S, Simon D, Zwicky C, Conus S, Simon HU. cIAP-2 and survivin contribute to cytokine-mediated delayed eosinophil apoptosis. Eur J Immunol. 2006;36:1975–84.
Hasegawa T, Suzuki K, Sakamoto C, Ohta K, Nishiki S, Hino M, et al. Expression of the inhibitor of apoptosis (IAP) family members in human neutrophils: up-regulation of cIAP2 by granulocyte colony-stimulating factor and overexpression of cIAP2 in chronic neutrophilic leukemia. Blood. 2003;101:1164–71.
Moulding DA, Quayle JA, Hart CA, Edwards SW. Mcl-1 expression in human neutrophils: regulation by cytokines and correlation with cell survival. Blood. 1998;92:2495–502.
Dibbert B, Daigle I, Braun D, Schranz C, Weber M, Blaser K, et al. Role for Bcl-xL in delayed eosinophil apoptosis mediated by granulocyte-macrophage colony-stimulating factor and interleukin-5. Blood. 1998;92:778–83.
Liew PX, Kubes P. The neutrophil’s role during health and disease. Physiol Rev. 2019;99:1223–48.
Arlet JB, Ribeil JA, Guillem F, Negre O, Hazoume A, Marcion G, et al. HSP70 sequestration by free α-globin promotes ineffective erythropoiesis in β-thalassaemia. Nature. 2014;514:242–6.
Gastou M, Rio S, Dussiot M, Karboul N, Moniz H, Leblanc T, et al. The severe phenotype of Diamond-Blackfan anemia is modulated by heat shock protein 70. Blood Adv. 2017;1:1959–76.
Duplomb L, Rivière J, Jego G, Da Costa R, Hammann A, Racine J, et al. Serpin B1 defect and increased apoptosis of neutrophils in Cohen syndrome neutropenia. J Mol Med. 2019;97:633–45.
Schwulst SJ, Muenzer JT, Peck-Palmer OM, Chang KC, Davis CG, McDonough JS, et al. Bim siRNA decreases lymphocyte apoptosis and improves survival in sepsis. Shock. 2008;30:127–34.
Chung CS, Venet F, Chen Y, Jones LN, Wilson DC, Ayala CA, et al. Deficiency of Bid protein reduces sepsis-induced apoptosis and inflammation, while improving septic survival. Shock. 2010;34:150–61.
Yan J, Zhang H, Xiang J, Zhao Y, Yuan X, Sun B, et al. The BH3-only protein BAD mediates TNFα cytotoxicity despite concurrent activation of IKK and NF-κB in septic shock. Cell Res. 2018;28:701–18.
Weber SU, Schewe JC, Lehmann LE, Müller S, Book M, Klaschik S, et al. Induction of Bim and Bid gene expression during accelerated apoptosis in severe sepsis. Crit Care. 2008;12:R128.
Oberholzer C, Tschoeke SK, Moldawer LL, Oberholzer A. Local thymic caspase-9 inhibition improves survival during polymicrobial sepsis in mice. J Mol Med. 2006;84:389–95.
Lamkanfi M, Moreira LO, Makena P, Spierings DC, Boyd K, Murray PJ, et al. Caspase-7 deficiency protects from endotoxin-induced lymphocyte apoptosis and improves survival. Blood. 2009;113:2742–5.
Yasuda T, Takeyama Y, Ueda T, Shinzeki M, Kishi S, Sawa H, et al. Protective effect of caspase inhibitor on intestinal integrity in experimental severe acute pancreatitis. J Surg Res. 2007;138:300–7.
Liu Y, Chen XD, Yu J, Chi JL, Long FW, Yang HW, et al. Deletion Of XIAP reduces the severity of acute pancreatitis via regulation of cell death and nuclear factor-κB activity. Cell Death Dis. 2017;8:e2685.
Leucht K, Caj M, Fried M, Rogler G, Hausmann M. Impaired removal of Vβ8(+) lymphocytes aggravates colitis in mice deficient for B cell lymphoma-2-interacting mediator of cell death (Bim). Clin Exp Immunol. 2013;173:493–501.
Wicki S, Gurzeler U, Corazza N, Genitsch V, Wong WW, Kaufmann T. Loss of BID delays FASL-induced cell death of mouse neutrophils and aggravates DSS-induced weight loss. Int J Mol Sci. 2018;19:684.
Weder B, Mozaffari M, Biedermann L, Mamie C, Moncsek A, Wang L, et al. BCL-2 levels do not predict azathioprine treatment response in inflammatory bowel disease, but inhibition induces lymphocyte apoptosis and ameliorates colitis in mice. Clin Exp Immunol. 2018;193:346–60.
Lutz C, Mozaffari M, Tosevski V, Caj M, Cippà P, McRae BL, et al. Increased lymphocyte apoptosis in mouse models of colitis upon ABT-737 treatment is dependent upon BIM expression. Clin Exp Immunol. 2015;181:343–56.
Dirisina R, Katzman RB, Goretsky T, Managlia E, Mittal N, Williams DB, et al. p53 and PUMA independently regulate apoptosis of intestinal epithelial cells in patients and mice with colitis. Gastroenterology. 2011;141:1036–45.
Qiu W, Carson-Walter EB, Liu H, Epperly M, Greenberger JS, Zambetti GP, et al. PUMA regulates intestinal progenitor cell radiosensitivity and gastrointestinal syndrome. cell Stem Cell. 2008;2:576–83.
Kirsch DG, Santiago PM, di Tomaso E, Sullivan JM, Hou WS, Dayton T, et al. p53 controls radiation-induced gastrointestinal syndrome in mice independent of apoptosis. Science. 2010;327:593–6.
Tan S, Wei X, Song M, Tao J, Yang Y, Khatoon S, et al. PUMA mediates ER stress-induced apoptosis in portal hypertensive gastropathy. Cell Death Dis. 2014;5:e1128.
Qiu W, Wu B, Wang X, Buchanan ME, Regueiro MD, Hartman DJ, et al. PUMA-mediated intestinal epithelial apoptosis contributes to ulcerative colitis in humans and mice. J Clin Investig. 2011;121:1722–32.
Wu B, Qiu W, Wang P, Yu H, Cheng T, Zambetti GP, et al. p53 independent induction of PUMA mediates intestinal apoptosis in response to ischaemia-reperfusion. Gut. 2007;56:645–54.
Coopersmith CM, O’Donnell D, Gordon JI. Bcl-2 inhibits ischemia-reperfusion-induced apoptosis in the intestinal epithelium of transgenic mice. Am J Physiol. 1999;276:G677–86.
Damgaard RB, Fiil BK, Speckmann C, Yabal M, zur Stadt U, Bekker-Jensen S, et al. Disease-causing mutations in the XIAP BIR2 domain impair NOD2-dependent immune signalling. EMBO Mol Med. 2013;5:1278–95.
Damgaard RB, Nachbur U, Yabal M, Wong WW, Fiil BK, Kastirr M, et al. The ubiquitin ligase XIAP recruits LUBAC for NOD2 signaling in inflammation and innate immunity. Mol Cell. 2012;46:746–58.
Yang X, Kanegane H, Nishida N, Imamura T, Hamamoto K, Miyashita R, et al. Clinical and genetic characteristics of XIAP deficiency in Japan. J Clin Immunol. 2012;32:411–20.
Salzer U, Hagena T, Webster DB, Grimbacher B. Sequence analysis of BIRC4/XIAP in male patients with common variable immunodeficiency. Int Arch Allergy Immunol. 2008;147:147–51.
Wahida A, Muller M, Hiergeist A, Popper B, Steiger K, Branca C, et al. XIAP restrains TNF-driven intestinal inflammation and dysbiosis by promoting innate immune responses of Paneth and dendritic cells. Sci Immunol. 2021;6:eabf7235.
Brinkman BM, Hildebrand F, Kubica M, Goosens D, Del Favero J, Declercq W, et al. Caspase deficiency alters the murine gut microbiome. Cell Death Dis. 2011;2:e220.
Ghazavi F, Huysentruyt J, De Coninck J, Kourula S, Martens S, Hassannia B, et al. Executioner caspases 3 and 7 are dispensable for intestinal epithelium turnover and homeostasis at steady state. Proc Natl Acad Sci USA. 2022;119:e2024508119.
Galluzzi L, Brenner C, Morselli E, Touat Z, Kroemer G. Viral control of mitochondrial apoptosis. PLoS Pathog. 2008;4:e1000018.
Günther SD, Fritsch M, Seeger JM, Schiffmann LM, Snipas SJ, Coutelle M, et al. Cytosolic Gram-negative bacteria prevent apoptosis by inhibition of effector caspases through lipopolysaccharide. Nat Microbiol. 2020;5:354–67.
Suzuki T, Okamoto T, Katoh H, Sugiyama Y, Kusakabe S, Tokunaga M, et al. Infection with flaviviruses requires BCLXL for cell survival. PLoS Pathog. 2018;14:e1007299.
Handke W, Luig C, Popovic B, Krmpotic A, Jonjic S, Brune W. Viral inhibition of BAK promotes murine cytomegalovirus dissemination to salivary glands. J Virol. 2013;87:3592–6.
Fleming P, Kvansakul M, Voigt V, Kile BT, Kluck RM, Huang DC, et al. MCMV-mediated inhibition of the pro-apoptotic Bak protein is required for optimal in vivo replication. PLoS Pathog. 2013;9:e1003192.
Garrison SP, Thornton JA, Häcker H, Webby R, Rehg JE, Parganas E, et al. The p53-target gene puma drives neutrophil-mediated protection against lethal bacterial sepsis. PLoS Pathog. 2010;6:e1001240.
Andree M, Seeger JM, Schüll S, Coutelle O, Wagner-Stippich D, Wiegmann K, et al. BID-dependent release of mitochondrial SMAC dampens XIAP-mediated immunity against Shigella. EMBO J. 2014;33:2171–87.
Stafford CA, Lawlor KE, Heim VJ, Bankovacki A, Bernardini JP, Silke J, et al. IAPs regulate distinct innate immune pathways to co-ordinate the response to bacterial peptidoglycans. Cell Rep. 2018;22:1496–508.
Margaroli C, Oberle S, Lavanchy C, Scherer S, Rosa M, Strasser A, et al. Role of proapoptotic BH3-only proteins in Listeria monocytogenes infection. Eur J Immunol. 2016;46:1427–37.
Bradfute SB, Swanson PE, Smith MA, Watanabe E, McDunn JE, Hotchkiss RS, et al. Mechanisms and consequences of ebolavirus-induced lymphocyte apoptosis. J Immunol. 2010;184:327–35.
Yang Y, Wu Y, Meng X, Wang Z, Younis M, Liu Y, et al. SARS-CoV-2 membrane protein causes the mitochondrial apoptosis and pulmonary edema via targeting BOK. Cell Death Differ. 2022;29:1395–408.
Anderson CJ, Medina CB, Barron BJ, Karvelyte L, Aaes TL, Lambertz I, et al. Microbes exploit death-induced nutrient release by gut epithelial cells. Nature. 2021;596:262–7.
Beckham JD, Tuttle KD, Tyler KL. Caspase-3 activation is required for reovirus-induced encephalitis in vivo. J Neurovirol. 2010;16:306–17.
Fischer SF, Belz GT, Strasser A. BH3-only protein Puma contributes to death of antigen-specific T cells during shutdown of an immune response to acute viral infection. Proc Natl Acad Sci USA. 2008;105:3035–40.
Villunger A, Michalak EM, Coultas L, Müllauer F, Böck G, Ausserlechner MJ, et al. p53- and drug-induced apoptotic responses mediated by BH3-only proteins puma and noxa. Science. 2003;302:1036–8.
Pellegrini M, Bouillet P, Robati M, Belz GT, Davey GM, Strasser A. Loss of Bim increases T cell production and function in interleukin 7 receptor-deficient mice. J Exp Med. 2004;200:1189–95.
Speir M, Lawlor KE, Glaser SP, Abraham G, Chow S, Vogrin A, et al. Eliminating Legionella by inhibiting BCL-XL to induce macrophage apoptosis. Nat Microbiol. 2016;1:15034.
Tam BT, Yu AP, Tam EW, Monks DA, Wang XP, Pei XM, et al. Ablation of Bax and Bak protects skeletal muscle against pressure-induced injury. Sci Rep. 2018;8:3689.
Stratos I, Li Z, Rotter R, Herlyn P, Mittlmeier T, Vollmar B. Inhibition of caspase mediated apoptosis restores muscle function after crush injury in rat skeletal muscle. Apoptosis. 2012;17:269–77.
Teng BT, Tam EW, Benzie IF, Siu PM. Protective effect of caspase inhibition on compression-induced muscle damage. J Physiol. 2011;58913:3349–69.
Talbert EE, Smuder AJ, Min K, Kwon OS, Powers SK. Calpain and caspase-3 play required roles in immobilization-induced limb muscle atrophy. J Appl Physiol. 2013;114:1482–9.
Zhu S, Nagashima M, Khan MA, Yasuhara S, Kaneki M, Martyn JA. Lack of caspase-3 attenuates immobilization-induced muscle atrophy and loss of tension generation along with mitigation of apoptosis and inflammation. Muscle Nerve. 2013;47:711–21.
Wang XH, Hu J, Du J, Klein JD. X-chromosome linked inhibitor of apoptosis protein inhibits muscle proteolysis in insulin-deficient mice. Gene Ther. 2007;14:711–20.
Hu J, Du J, Zhang L, Price SR, Klein JD, Wang XH. XIAP reduces muscle proteolysis induced by CKD. J Am Soc Nephrol. 2010;21:1174–83.
Plant PJ, Bain JR, Correa JE, Woo M, Batt J. Absence of caspase-3 protects against denervation-induced skeletal muscle atrophy. J Appl Physiol. 2009;107:224–34.
Zhu H, Pytel P, Gomez CM. Selective inhibition of caspases in skeletal muscle reverses the apoptotic synaptic degeneration in slow-channel myasthenic syndrome. Hum Mol Genet. 2014;23:69–77.
Budinger GR, Mutlu GM, Urich D, Soberanes S, Buccellato LJ, Hawkins K, et al. Epithelial cell death is an important contributor to oxidant-mediated acute lung injury. Am J Respir Crit Care Med. 2011;183:1043–54.
He CH, Waxman AB, Lee CG, Link H, Rabach ME, Ma B, et al. Bcl-2-related protein A1 is an endogenous and cytokine-stimulated mediator of cytoprotection in hyperoxic acute lung injury. J Clin Investig. 2005;115:1039–48.
Métrailler-Ruchonnet I, Pagano A, Carnesecchi S, Khatib K, Herrera P, Donati Y, et al. Bcl-2 overexpression in type II epithelial cells does not prevent hyperoxia-induced acute lung injury in mice. Am J Physiol Lung Cell Mol Physiol. 2010;299:L312–322.
Gangoda L, Schenk RL, Best SA, Nedeva C, Louis C, D’Silva DB, et al. Absence of pro-survival A1 has no impact on inflammatory cell survival in vivo during acute lung inflammation and peritonitis. Cell Death Differ. 2022;29:96–104.
Kang HR, Cho SJ, Lee CG, Homer RJ, Elias JA. Transforming growth factor (TGF)-beta1 stimulates pulmonary fibrosis and inflammation via a Bax-dependent, bid-activated pathway that involves matrix metalloproteinase-12. J Biol Chem. 2007;282:7723–32.
Budinger GR, Mutlu GM, Eisenbart J, Fuller AC, Bellmeyer AA, Baker CM, et al. Proapoptotic Bid is required for pulmonary fibrosis. Proc Natl Acad Sci USA. 2006;103:4604–9.
Gu L, Surolia R, Larson-Casey JL, He C, Davis D, Kang J, et al. Targeting Cpt1a-Bcl-2 interaction modulates apoptosis resistance and fibrotic remodeling. Cell Death Differ. 2022;29:118–32.
Kuwano K, Kunitake R, Maeyama T, Hagimoto N, Kawasaki M, Matsuba T, et al. Attenuation of bleomycin-induced pneumopathy in mice by a caspase inhibitor. Am J Physiol Lung Cell Mol Physiol. 2001;280:L316–25.
Wang R, Ibarra-Sunga O, Verlinski L, Pick R, Uhal BD. Abrogation of bleomycin-induced epithelial apoptosis and lung fibrosis by captopril or by a caspase inhibitor. Am J Physiol Lung Cell Mol Physiol. 2000;279:L143–51.
Wang HL, Akinci IO, Baker CM, Urich D, Bellmeyer A, Jain M, et al. The intrinsic apoptotic pathway is required for lipopolysaccharide-induced lung endothelial cell death. J Immunol. 2007;179:1834–41.
Zhang YX, Fan H, Shi Y, Xu ST, Yuan YF, Zheng RH, et al. Prevention of lung ischemia-reperfusion injury by short hairpin RNA-mediated caspase-3 gene silencing. J Thorac Cardiovasc Surg. 2010;139:758–64.
Wang L, Chen B, Xiong X, Chen S, Jin L, Zhu M. Necrostatin-1 synergizes the Pan caspase inhibitor to attenuate lung injury induced by ischemia reperfusion in rats. Mediators Inflamm. 2020;2020:7059304.
Cooke DT, Hoyt EG, Robbins RC. Overexpression of human Bcl-2 in syngeneic rat donor lungs preserves posttransplant function and reduces intragraft caspase activity and interleukin-1beta production. Transplantation. 2005;79:762–7.
Quadri SM, Segall L, de Perrot M, Han B, Edwards V, Jones N, et al. Caspase inhibition improves ischemia-reperfusion injury after lung transplantation. Am J Transplant. 2005;5:292–9.
Liu M, Shi L, Zou X, Zheng X, Zhang F, Ding X, et al. Caspase inhibitor zVAD-fmk protects against acute pancreatitis-associated lung injury via inhibiting inflammation and apoptosis. Pancreatology. 2016;16:733–8.
Kawasaki M, Kuwano K, Hagimoto N, Matsuba T, Kunitake R, Tanaka T, et al. Protection from lethal apoptosis in lipopolysaccharide-induced acute lung injury in mice by a caspase inhibitor. Am J Pathol. 2000;157:597–603.
van den Berg E, Bal SM, Kuipers MT, Matute-Bello G, Lutter R, Bos AP, et al. The caspase inhibitor zVAD increases lung inflammation in pneumovirus infection in mice. Physiol Rep. 2015;3:e12332.
Locatelli F, Corti S, Papadimitriou D, Fortunato F, Del Bo R, Donadoni C, et al. Fas small interfering RNA reduces motoneuron death in amyotrophic lateral sclerosis mice. Ann Neurol. 2007;62:81–92.
Petri S, Kiaei M, Wille E, Calingasan NY, Flint Beal M. Loss of Fas ligand-function improves survival in G93A-transgenic ALS mice. J Neurol Sci. 2006;251:44–49.
Gowing G, Dequen F, Soucy G, Julien JP. Absence of tumor necrosis factor-alpha does not affect motor neuron disease caused by superoxide dismutase 1 mutations. J Neurosci. 2006;26:11397–402.
Tortarolo M, Vallarola A, Lidonnici D, Battaglia E, Gensano F, Spaltro G, et al. Lack of TNF-alpha receptor type 2 protects motor neurons in a cellular model of amyotrophic lateral sclerosis and in mutant SOD1 mice but does not affect disease progression. J Neurochem. 2015;135:109–24.
Bartsch JW, Wildeboer D, Koller G, Naus S, Rittger A, Moss ML, et al. Tumor necrosis factor-alpha (TNF-alpha) regulates shedding of TNF-alpha receptor 1 by the metalloprotease-disintegrin ADAM8: evidence for a protease-regulated feedback loop in neuroprotection. J Neurosci. 2010;30:12210–8.
Bernard-Marissal N, Moumen A, Sunyach C, Pellegrino C, Dudley K, Henderson CE, et al. Reduced calreticulin levels link endoplasmic reticulum stress and Fas-triggered cell death in motoneurons vulnerable to ALS. J Neurosci. 2012;32:4901–12.
Guégan C, Vila M, Teismann P, Chen C, Onténiente B, Li M, et al. Instrumental activation of bid by caspase-1 in a transgenic mouse model of ALS. Mol Cell Neurosci. 2002;20:553–62.
Kalovyrna N, Apokotou O, Boulekou S, Paouri E, Boutou A, Georgopoulos S. A 3’UTR modification of the TNF-α mouse gene increases peripheral TNF-α and modulates the Alzheimer-like phenotype in 5XFAD mice. Sci Rep. 2020;10:8670.
Paouri E, Tzara O, Zenelak S, Georgopoulos S. Genetic deletion of tumor necrosis factor-α attenuates amyloid-β production and decreases amyloid plaque formation and glial response in the 5XFAD model of Alzheimer’s disease. J Alzheimer’s Dis. 2017;60:165–81.
Paouri E, Tzara O, Kartalou GI, Zenelak S, Georgopoulos S. Peripheral tumor necrosis factor-alpha (TNF-α) modulates amyloid pathology by regulating blood-derived immune cells and glial response in the brain of AD/TNF Transgenic Mice. J Neurosci. 2017;37:5155–71.
Tweedie D, Ferguson RA, Fishman K, Frankola KA, Van Praag H, Holloway HW, et al. Tumor necrosis factor-α synthesis inhibitor 3,6’-dithiothalidomide attenuates markers of inflammation, Alzheimer pathology and behavioral deficits in animal models of neuroinflammation and Alzheimer’s disease. J Neuroinflammation. 2012;9:106.
McAlpine FE, Lee JK, Harms AS, Ruhn KA, Blurton-Jones M, Hong J, et al. Inhibition of soluble TNF signaling in a mouse model of Alzheimer’s disease prevents pre-plaque amyloid-associated neuropathology. Neurobiol Dis. 2009;34:163–77.
MacPherson KP, Sompol P, Kannarkat GT, Chang J, Sniffen L, Wildner ME, et al. Peripheral administration of the soluble TNF inhibitor XPro1595 modifies brain immune cell profiles, decreases beta-amyloid plaque load, and rescues impaired long-term potentiation in 5xFAD mice. Neurobiol Dis. 2017;102:81–95.
Gabbita SP, Johnson MF, Kobritz N, Eslami P, Poteshkina A, Varadarajan S, et al. Oral TNFα modulation alters neutrophil infiltration, improves cognition and diminishes tau and amyloid pathology in the 3xTgAD mouse model. PLoS ONE. 2015;10:e0137305.
Gabbita SP, Srivastava MK, Eslami P, Johnson MF, Kobritz NK, Tweedie D, et al. Early intervention with a small molecule inhibitor for tumor necrosis factor-α prevents cognitive deficits in a triple transgenic mouse model of Alzheimer’s disease. J Neuroinflammation. 2012;9:99.
Lourenco MV, Clarke JR, Frozza RL, Bomfim TR, Forny-Germano L, Batista AF, et al. TNF-α mediates PKR-dependent memory impairment and brain IRS-1 inhibition induced by Alzheimer’s β-amyloid oligomers in mice and monkeys. Cell Metab. 2013;18:831–43.
Camargos S, Scholz S, Simón-Sánchez J, Paisán-Ruiz C, Lewis P, Hernandez D, et al. DYT16, a novel young-onset dystonia-parkinsonism disorder: identification of a segregating mutation in the stress-response protein PRKRA. Lancet Neurol. 2008;7:207–15.
Bhaskar K, Maphis N, Xu G, Varvel NH, Kokiko-Cochran ON, Weick JP, et al. Microglial derived tumor necrosis factor-α drives Alzheimer’s disease-related neuronal cell cycle events. Neurobiol Dis. 2014;62:273–85.
Steeland S, Gorlé N, Vandendriessche C, Balusu S, Brkic M, Van Cauwenberghe C, et al. Counteracting the effects of TNF receptor-1 has therapeutic potential in Alzheimer’s disease. EMBO Mol Med. 2018;10:e8300.
He P, Zhong Z, Lindholm K, Berning L, Lee W, Lemere C, et al. Deletion of tumor necrosis factor death receptor inhibits amyloid beta generation and prevents learning and memory deficits in Alzheimer’s mice. J Cell Biol. 2007;178:829–41.
Jayaraman A, Htike TT, James R, Picon C, Reynolds R. TNF-mediated neuroinflammation is linked to neuronal necroptosis in Alzheimer’s disease hippocampus. Acta Neuropathol Commun. 2021;9:159.
Xu C, Wu J, Wu Y, Ren Z, Yao Y, Chen G, et al. TNF-alpha-dependent neuronal necroptosis regulated in Alzheimer’s disease by coordination of RIPK1-p62 complex with autophagic UVRAG. Theranostics. 2021;11:9452–69.
Montgomery SL, Mastrangelo MA, Habib D, Narrow WC, Knowlden SA, Wright TW, et al. Ablation of TNF-RI/RII expression in Alzheimer’s disease mice leads to an unexpected enhancement of pathology: implications for chronic pan-TNF-α suppressive therapeutic strategies in the brain. Am J Pathol. 2011;179:2053–70.
Montgomery SL, Narrow WC, Mastrangelo MA, Olschowka JA, O’Banion MK, Bowers WJ. Chronic neuron- and age-selective down-regulation of TNF receptor expression in triple-transgenic Alzheimer disease mice leads to significant modulation of amyloid- and Tau-related pathologies. Am J Pathol. 2013;182:2285–97.
Ferger B, Leng A, Mura A, Hengerer B, Feldon J. Genetic ablation of tumor necrosis factor-alpha (TNF-alpha) and pharmacological inhibition of TNF-synthesis attenuates MPTP toxicity in mouse striatum. J Neurochem. 2004;89:822–33.
Sriram K, Matheson JM, Benkovic SA, Miller DB, Luster MI, O’Callaghan JP. Mice deficient in TNF receptors are protected against dopaminergic neurotoxicity: implications for Parkinson’s disease. FASEB J. 2002;16:1474–6.
Zhou QH, Sumbria R, Hui EK, Lu JZ, Boado RJ, Pardridge WM. Neuroprotection with a brain-penetrating biologic tumor necrosis factor inhibitor. J Pharmacol Exp Ther. 2011;339:618–23.
McCoy MK, Martinez TN, Ruhn KA, Szymkowski DE, Smith CG, Botterman BR, et al. Blocking soluble tumor necrosis factor signaling with dominant-negative tumor necrosis factor inhibitor attenuates loss of dopaminergic neurons in models of Parkinson’s disease. J Neurosci. 2006;26:9365–75.
Sriram K, Matheson JM, Benkovic SA, Miller DB, Luster MI, O’Callaghan JP. Deficiency of TNF receptors suppresses microglial activation and alters the susceptibility of brain regions to MPTP-induced neurotoxicity: role of TNF-alpha. FASEB J. 2006;20:670–82.
Dong Y, Fischer R, Naudé PJ, Maier O, Nyakas C, Duffey M, et al. Essential protective role of tumor necrosis factor receptor 2 in neurodegeneration. Proc Natl Acad Sci USA. 2016;113:12304–9.
Shi JQ, Wang BR, Jiang WW, Chen J, Zhu YW, Zhong LL, et al. Cognitive improvement with intrathecal administration of infliximab in a woman with Alzheimer’s disease. J Am Geriatr Soc. 2011;59:1142–4.
Tobinick EL, Gross H. Rapid cognitive improvement in Alzheimer’s disease following perispinal etanercept administration. J Neuroinflammation. 2008;5:2.
Alto LT, Chen X, Ruhn KA, Treviño I, Tansey MG. AAV-dominant negative tumor necrosis factor (DN-TNF) gene transfer to the striatum does not rescue medium spiny neurons in the YAC128 mouse model of Huntington’s disease. PLoS ONE. 2014;9:e96544.
Cantarella G, Di Benedetto G, Puzzo D, Privitera L, Loreto C, Saccone S, et al. Neutralization of TNFSF10 ameliorates functional outcome in a murine model of Alzheimer’s disease. Brain. 2015;138:203–16.
Uberti D, Ferrari-Toninelli G, Bonini SA, Sarnico I, Benarese M, Pizzi M, et al. Blockade of the tumor necrosis factor-related apoptosis inducing ligand death receptor DR5 prevents beta-amyloid neurotoxicity. Neuropsychopharmacology. 2007;32:872–80.
Takahashi T, Tanaka M, Brannan CI, Jenkins NA, Copeland NG, Suda T, et al. Generalized lymphoproliferative disease in mice, caused by a point mutation in the Fas ligand. Cell. 1994;76:969–76.
Landau AM, Luk KC, Jones ML, Siegrist-Johnstone R, Young YK, Kouassi E, et al. Defective Fas expression exacerbates neurotoxicity in a model of Parkinson’s disease. J Exp Med. 2005;202:575–81.
Gao L, Brenner D, Llorens-Bobadilla E, Saiz-Castro G, Frank T, Wieghofer P, et al. Infiltration of circulating myeloid cells through CD95L contributes to neurodegeneration in mice. J Exp Med. 2015;212:469–80.
Hayley S, Crocker SJ, Smith PD, Shree T, Jackson-Lewis V, Przedborski S, et al. Regulation of dopaminergic loss by Fas in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s disease. J Neurosci. 2004;24:2045–53.
Betarbet R, Anderson LR, Gearing M, Hodges TR, Fritz JJ, Lah JJ, et al. Fas-associated factor 1 and Parkinson’s disease. Neurobiol Dis. 2008;31:309–15.
Sul JW, Park MY, Shin J, Kim YR, Yoo SE, Kong YY, et al. Accumulation of the parkin substrate, FAF1, plays a key role in the dopaminergic neurodegeneration. Hum Mol Genet. 2013;22:1558–73.
Rohn TT, Head E, Nesse WH, Cotman CW, Cribbs DH. Activation of caspase-8 in the Alzheimer’s disease brain. Neurobiol Dis. 2001;8:1006–16.
Sánchez I, Xu CJ, Juo P, Kakizaka A, Blenis J, Yuan J. Caspase-8 is required for cell death induced by expanded polyglutamine repeats. Neuron. 1999;22:623–33.
Yang L, Sugama S, Mischak RP, Kiaei M, Bizat N, Brouillet E, et al. A novel systemically active caspase inhibitor attenuates the toxicities of MPTP, malonate, and 3NP in vivo. Neurobiol Dis. 2004;17:250–9.
Viceconte N, Burguillos MA, Herrera AJ, De Pablos RM, Joseph B, Venero JL. Neuromelanin activates proinflammatory microglia through a caspase-8-dependent mechanism. J Neuroinflammation. 2015;12:5.
Fricker M, Vilalta A, Tolkovsky AM, Brown GC. Caspase inhibitors protect neurons by enabling selective necroptosis of inflamed microglia. J Biol Chem. 2013;288:9145–52.
Burguillos MA, Deierborg T, Kavanagh E, Persson A, Hajji N, Garcia-Quintanilla A, et al. Caspase signalling controls microglia activation and neurotoxicity. Nature. 2011;472:319–24.
Kavanagh E, Burguillos MA, Carrillo-Jimenez A, Oliva-Martin MJ, Santiago M, Rodhe J, et al. Deletion of caspase-8 in mouse myeloid cells blocks microglia pro-inflammatory activation and confers protection in MPTP neurodegeneration model. Aging. 2015;7:673–89.
Xu D, Zhao H, Jin M, Zhu H, Shan B, Geng J, et al. Modulating TRADD to restore cellular homeostasis and inhibit apoptosis. Nature. 2020;587:133–8.
Hartmann A, Troadec JD, Hunot S, Kikly K, Faucheux BA, Mouatt-Prigent A, et al. Caspase-8 is an effector in apoptotic death of dopaminergic neurons in Parkinson’s disease, but pathway inhibition results in neuronal necrosis. J Neurosci. 2001;21:2247–55.
Rehker J, Rodhe J, Nesbitt RR, Boyle EA, Martin BK, Lord J, et al. Caspase-8, association with Alzheimer’s Disease and functional analysis of rare variants. PLoS ONE. 2017;12:e0185777.
Jasmin M, Ahn EH, Voutilainen MH, Fombonne J, Guix C, Viljakainen T, et al. Netrin-1 and its receptor DCC modulate survival and death of dopamine neurons and Parkinson’s disease features. EMBO J. 2021;40:e105537.
Yu WR, Fehlings MG. Fas/FasL-mediated apoptosis and inflammation are key features of acute human spinal cord injury: implications for translational, clinical application. Acta Neuropathol. 2011;122:747–61.
Casha S, Yu WR, Fehlings MG. FAS deficiency reduces apoptosis, spares axons and improves function after spinal cord injury. Exp Neurol. 2005;196:390–400.
Demjen D, Klussmann S, Kleber S, Zuliani C, Stieltjes B, Metzger C, et al. Neutralization of CD95 ligand promotes regeneration and functional recovery after spinal cord injury. Nat Med. 2004;10:389–95.
Yu WR, Liu T, Fehlings TK, Fehlings MG. Involvement of mitochondrial signaling pathways in the mechanism of Fas-mediated apoptosis after spinal cord injury. Eur J Neurosci. 2009;29:114–31.
Letellier E, Kumar S, Sancho-Martinez I, Krauth S, Funke-Kaiser A, Laudenklos S, et al. CD95-ligand on peripheral myeloid cells activates Syk kinase to trigger their recruitment to the inflammatory site. Immunity. 2010;32:240–52.
Ellman DG, Lund MC, Nissen M, Nielsen PS, Sørensen C, Lester EB, et al. Conditional ablation of myeloid TNF improves functional outcome and decreases lesion size after spinal cord injury in mice. Cells. 2020;9.
Ziebell JM, Bye N, Semple BD, Kossmann T, Morganti-Kossmann MC. Attenuated neurological deficit, cell death and lesion volume in Fas-mutant mice is associated with altered neuroinflammation following traumatic brain injury. Brain Res. 2011;1414:94–105.
Yu WR, Liu T, Kiehl TR, Fehlings MG. Human neuropathological and animal model evidence supporting a role for Fas-mediated apoptosis and inflammation in cervical spondylotic myelopathy. Brain. 2011;134:1277–92.
Yang J, You Z, Kim HH, Hwang SK, Khuman J, Guo S, et al. Genetic analysis of the role of tumor necrosis factor receptors in functional outcome after traumatic brain injury in mice. J Neurotrauma. 2010;27:1037–46.
Bermpohl D, You Z, Lo EH, Kim HH, Whalen MJ. TNF alpha and Fas mediate tissue damage and functional outcome after traumatic brain injury in mice. J Cereb Blood Flow Metab. 2007;27:1806–18.
Longhi L, Perego C, Ortolano F, Aresi S, Fumagalli S, Zanier ER, et al. Tumor necrosis factor in traumatic brain injury: effects of genetic deletion of p55 or p75 receptor. J Cereb Blood Flow Metab. 2013;33:1182–9.
Khuman J, Meehan WP 3rd, Zhu X, Qiu J, Hoffmann U, Zhang J, et al. Tumor necrosis factor alpha and Fas receptor contribute to cognitive deficits independent of cell death after concussive traumatic brain injury in mice. J Cereb Blood Flow Metab. 2011;31:778–89.
Quintana A, Giralt M, Rojas S, Penkowa M, Campbell IL, Hidalgo J, et al. Differential role of tumor necrosis factor receptors in mouse brain inflammatory responses in cryolesion brain injury. J Neurosci Res. 2005;82:701–16.
Mironets E, Osei-Owusu P, Bracchi-Ricard V, Fischer R, Owens EA, Ricard J, et al. Soluble TNFα signaling within the spinal cord contributes to the development of autonomic dysreflexia and ensuing vascular and immune dysfunction after spinal cord injury. J Neurosci. 2018;38:4146–62.
Baratz R, Tweedie D, Wang JY, Rubovitch V, Luo W, Hoffer BJ, et al. Transiently lowering tumor necrosis factor-α synthesis ameliorates neuronal cell loss and cognitive impairments induced by minimal traumatic brain injury in mice. J Neuroinflammation. 2015;12:45.
Chen KB, Uchida K, Nakajima H, Yayama T, Hirai T, Watanabe S, et al. Tumor necrosis factor-α antagonist reduces apoptosis of neurons and oligodendroglia in rat spinal cord injury. Spine. 2011;36:1350–8.
O’Reilly ML, Mironets E, Shapiro TM, Crowther K, Collyer E, Bethea JR, et al. Pharmacological inhibition of soluble tumor necrosis factor-alpha two weeks after high thoracic spinal cord injury does not affect sympathetic hyperreflexia. J Neurotrauma. 2021;38:2186–91.
Ellman DG, Degn M, Lund MC, Clausen BH, Novrup HG, Flæng SB, et al. Genetic ablation of soluble TNF does not affect lesion size and functional recovery after moderate spinal cord injury in mice. Mediators Inflamm. 2016;2016:2684098.
Oshima T, Lee S, Sato A, Oda S, Hirasawa H, Yamashita T. TNF-alpha contributes to axonal sprouting and functional recovery following traumatic brain injury. Brain Res. 2009;1290:102–10.
Kim GM, Xu J, Xu J, Song SK, Yan P, Ku G, et al. Tumor necrosis factor receptor deletion reduces nuclear factor-kappaB activation, cellular inhibitor of apoptosis protein 2 expression, and functional recovery after traumatic spinal cord injury. J Neurosci. 2001;21:6617–25.
Scherbel U, Raghupathi R, Nakamura M, Saatman KE, Trojanowski JQ, Neugebauer E, et al. Differential acute and chronic responses of tumor necrosis factor-deficient mice to experimental brain injury. Proc Natl Acad Sci USA. 1999;96:8721–6.
Cantarella G, Di Benedetto G, Scollo M, Paterniti I, Cuzzocrea S, Bosco P, et al. Neutralization of tumor necrosis factor-related apoptosis-inducing ligand reduces spinal cord injury damage in mice. Neuropsychopharmacol. 2010;35:1302–14.
Fang Y, Lu J, Wang X, Wu H, Mei S, Zheng J, et al. HIF-1α mediates TRAIL-Induced Neuronal Apoptosis via regulating DcR1 expression following traumatic brain injury. Front Cell Neurosci. 2020;14:192.
Sobrido-Cameán D, Barreiro-Iglesias A. Role of Caspase-8 and Fas in cell death after spinal cord injury. Front Mol Neurosci. 2018;11:101.
Sung TC, Chen Z, Thuret S, Vilar M, Gage FH, Riek R, et al. P45 forms a complex with FADD and promotes neuronal cell survival following spinal cord injury. PLoS ONE. 2013;8:e69286.
Krajewska M, You Z, Rong J, Kress C, Huang X, Yang J, et al. Neuronal deletion of caspase 8 protects against brain injury in mouse models of controlled cortical impact and kainic acid-induced excitotoxicity. PLoS ONE. 2011;6:e24341.
Ugolini G, Raoul C, Ferri A, Haenggeli C, Yamamoto Y, Salaün D, et al. Fas/tumor necrosis factor receptor death signaling is required for axotomy-induced death of motoneurons in vivo. J Neurosci. 2003;23:8526–31.
Monnier PP, D’Onofrio PM, Magharious M, Hollander AC, Tassew N, Szydlowska K, et al. Involvement of caspase-6 and caspase-8 in neuronal apoptosis and the regenerative failure of injured retinal ganglion cells. J Neurosci. 2011;31:10494–505.
Tezel G, Yang X, Yang J, Wax MB. Role of tumor necrosis factor receptor-1 in the death of retinal ganglion cells following optic nerve crush injury in mice. Brain Res. 2004;996:202–12.
Fontaine V, Mohand-Said S, Hanoteau N, Fuchs C, Pfizenmaier K, Eisel U. Neurodegenerative and neuroprotective effects of tumor Necrosis factor (TNF) in retinal ischemia: opposite roles of TNF receptor 1 and TNF receptor 2. J Neurosci. 2002;22:Rc216.
Nakazawa T, Nakazawa C, Matsubara A, Noda K, Hisatomi T, She H, et al. Tumor necrosis factor-alpha mediates oligodendrocyte death and delayed retinal ganglion cell loss in a mouse model of glaucoma. J Neurosci. 2006;26:12633–41.
Krishnan A, Kocab AJ, Zacks DN, Marshak-Rothstein A, Gregory-Ksander M. A small peptide antagonist of the Fas receptor inhibits neuroinflammation and prevents axon degeneration and retinal ganglion cell death in an inducible mouse model of glaucoma. J Neuroinflammation. 2019;16:184.
Cueva Vargas JL, Osswald IK, Unsain N, Aurousseau MR, Barker PA, Bowie D, et al. Soluble tumor necrosis factor alpha promotes retinal ganglion cell death in glaucoma via calcium-permeable AMPA receptor activation. J Neurosci. 2015;35:12088–102.
Roh M, Zhang Y, Murakami Y, Thanos A, Lee SC, Vavvas DG, et al. Etanercept, a widely used inhibitor of tumor necrosis factor-α (TNF-α), prevents retinal ganglion cell loss in a rat model of glaucoma. PLoS ONE. 2012;7:e40065.
Yang X, Zeng Q, Tezel G. Regulation of distinct caspase-8 functions in retinal ganglion cells and astroglia in experimental glaucoma. Neurobiol Dis. 2021;150:105258.
Tisch N, Freire-Valls A, Yerbes R, Paredes I, La Porta S, Wang X, et al. Caspase-8 modulates physiological and pathological angiogenesis during retina development. J Clin Investig. 2019;129:5092–107.
Kang TB, Ben-Moshe T, Varfolomeev EE, Pewzner-Jung Y, Yogev N, Jurewicz A, et al. Caspase-8 serves both apoptotic and nonapoptotic roles. J Immunol. 2004;173:2976–84.
Tian Y, Li H, Liu X, Xie L, Huang Z, Li W, et al. Pharmacological inhibition of caspase-8 suppresses inflammation-induced angiogenesis in the cornea. Biomolecules. 2020;10:210.
Burgaletto C, Platania CBM, Di Benedetto G, Munafò A, Giurdanella G, Federico C, et al. Targeting the miRNA-155/TNFSF10 network restrains inflammatory response in the retina in a mouse model of Alzheimer’s disease. Cell Death Dis. 2021;12:905.
Meng HL, Li XX, Chen YT, Yu LJ, Zhang H, Lao JM, et al. Neuronal soluble fas ligand drives M1-microglia polarization after cerebral ischemia. CNS Neurosci Ther. 2016;22:771–81.
Niu FN, Zhang X, Hu XM, Chen J, Chang LL, Li JW, et al. Targeted mutation of Fas ligand gene attenuates brain inflammation in experimental stroke. Brain Behav Immun. 2012;26:61–71.
Martin-Villalba A, Herr I, Jeremias I, Hahne M, Brandt R, Vogel J, et al. CD95 ligand (Fas-L/APO-1L) and tumor necrosis factor-related apoptosis-inducing ligand mediate ischemia-induced apoptosis in neurons. J Neurosci. 1999;19:3809–17.
Graham EM, Sheldon RA, Flock DL, Ferriero DM, Martin LJ, O’Riordan DP, et al. Neonatal mice lacking functional Fas death receptors are resistant to hypoxic-ischemic brain injury. Neurobiol Dis. 2004;17:89–98.
Dzietko M, Boos V, Sifringer M, Polley O, Gerstner B, Genz K, et al. A critical role for Fas/CD-95 dependent signaling pathways in the pathogenesis of hyperoxia-induced brain injury. Ann Neurol. 2008;64:664–73.
Ullah I, Chung K, Oh J, Beloor J, Bae S, Lee SC, et al. Intranasal delivery of a Fas-blocking peptide attenuates Fas-mediated apoptosis in brain ischemia. Sci Rep. 2018;8:15041.
Martin-Villalba A, Hahne M, Kleber S, Vogel J, Falk W, Schenkel J, et al. Therapeutic neutralization of CD95-ligand and TNF attenuates brain damage in stroke. Cell Death Differ. 2001;8:679–86.
Xu W, Jin W, Zhang X, Chen J, Ren C. Remote limb preconditioning generates a neuroprotective effect by modulating the extrinsic apoptotic pathway and TRAIL-receptors expression. Cell Mol Neurobiol. 2017;37:169–82.
Cui M, Wang L, Liang X, Ma X, Liu Y, Yang M, et al. Blocking TRAIL-DR5 signaling with soluble DR5 reduces delayed neuronal damage after transient global cerebral ischemia. Neurobiol Dis. 2010;39:138–47.
Clausen BH, Degn M, Sivasaravanaparan M, Fogtmann T, Andersen MG, Trojanowsky MD, et al. Conditional ablation of myeloid TNF increases lesion volume after experimental stroke in mice, possibly via altered ERK1/2 signaling. Sci Rep. 2016;6:29291.
Lambertsen KL, Clausen BH, Babcock AA, Gregersen R, Fenger C, Nielsen HH, et al. Microglia protect neurons against ischemia by synthesis of tumor necrosis factor. J Neurosci. 2009;29:1319–30.
Murakami Y, Saito K, Hara A, Zhu Y, Sudo K, Niwa M, et al. Increases in tumor necrosis factor-alpha following transient global cerebral ischemia do not contribute to neuron death in mouse hippocampus. J Neurochem. 2005;93:1616–22.
Bruce AJ, Boling W, Kindy MS, Peschon J, Kraemer PJ, Carpenter MK, et al. Altered neuronal and microglial responses to excitotoxic and ischemic brain injury in mice lacking TNF receptors. Nat Med. 1996;2:788–94.
Lei B, Dawson HN, Roulhac-Wilson B, Wang H, Laskowitz DT, James ML. Tumor necrosis factor α antagonism improves neurological recovery in murine intracerebral hemorrhage. J Neuroinflamm. 2013;10:103.
Yli-Karjanmaa M, Clausen BH, Degn M, Novrup HG, Ellman DG, Toft-Jensen P, et al. Topical administration of a soluble TNF inhibitor reduces infarct volume after focal cerebral ischemia in mice. Front Neurosci. 2019;13:781.
Madsen PM, Clausen BH, Degn M, Thyssen S, Kristensen LK, Svensson M, et al. Genetic ablation of soluble tumor necrosis factor with preservation of membrane tumor necrosis factor is associated with neuroprotection after focal cerebral ischemia. J Cereb Blood Flow Metab. 2016;36:1553–69.
Wu MH, Huang CC, Chio CC, Tsai KJ, Chang CP, Lin NK, et al. Inhibition of peripheral TNF-α and downregulation of microglial activation by alpha-lipoic acid and etanercept protect rat brain against ischemic stroke. Mol Neurobiol. 2016;53:4961–71.
Clausen BH, Degn M, Martin NA, Couch Y, Karimi L, Ormhøj M, et al. Systemically administered anti-TNF therapy ameliorates functional outcomes after focal cerebral ischemia. J Neuroinflammation. 2014;11:203.
Arango-Dávila CA, Vera A, Londoño AC, Echeverri AF, Cañas F, Cardozo CF, et al. Soluble or soluble/membrane TNF-α inhibitors protect the brain from focal ischemic injury in rats. Int J Neurosci. 2015;125:936–40.
Lu YM, Huang JY, Wang H, Lou XF, Liao MH, Hong LJ, et al. Targeted therapy of brain ischaemia using Fas ligand antibody conjugated PEG-lipid nanoparticles. Biomaterials. 2014;35:530–7.
Nawashiro H, Tasaki K, Ruetzler CA, Hallenbeck JM. TNF-alpha pretreatment induces protective effects against focal cerebral ischemia in mice. J Cereb Blood Flow Metab. 1997;17:483–90.
Kanazawa T, Kurano T, Ibaraki H, Takashima Y, Suzuki T, Seta Y. Therapeutic effects in a transient middle cerebral artery occlusion rat model by nose-to-brain delivery of anti-TNF-alpha siRNA with cell-penetrating peptide-modified polymer micelles. Pharmaceutics. 2019;11:478.
Lin SY, Wang YY, Chang CY, Wu CC, Chen WY, Liao SL, et al. TNF-α receptor inhibitor alleviates metabolic and inflammatory changes in a rat model of ischemic stroke. Antioxidants. 2021;10:851.
Xiaohong W, Jun Z, Hongmei G, Fan Q. CFLAR is a critical regulator of cerebral ischaemia-reperfusion injury through regulating inflammation and endoplasmic reticulum (ER) stress. Biomed Pharmacother. 2019;117:109155.
Taoufik E, Valable S, Müller GJ, Roberts ML, Divoux D, Tinel A, et al. FLIP(L) protects neurons against in vivo ischemia and in vitro glucose deprivation-induced cell death. J Neurosci. 2007;27:6633–46.
Ke DQ, Chen ZY, Li ZL, Huang X, Liang H. Target inhibition of caspase-8 alleviates brain damage after subarachnoid hemorrhage. Neural Regen Res. 2020;15:1283–9.
Shabanzadeh AP, D’Onofrio PM, Monnier PP, Koeberle PD. Targeting caspase-6 and caspase-8 to promote neuronal survival following ischemic stroke. Cell Death Dis. 2015;6:e1967.
Inoue S, Davis DP, Drummond JC, Cole DJ, Patel PM. The combination of isoflurane and caspase 8 inhibition results in sustained neuroprotection in rats subject to focal cerebral ischemia. Anesth Analg. 2006;102:1548–55.
Muhammad IF, Borné Y, Melander O, Orho-Melander M, Nilsson J, Söderholm M, et al. FADD (Fas-associated protein with death domain), caspase-3, and caspase-8 and incidence of ischemic stroke. Stroke. 2018;49:2224–6.
Rodhe J, Burguillos MA, de Pablos RM, Kavanagh E, Persson A, Englund E, et al. Spatio-temporal activation of caspase-8 in myeloid cells upon ischemic stroke. Acta Neuropathol Commun. 2016;4:92.
Taoufik E, Petit E, Divoux D, Tseveleki V, Mengozzi M, Roberts ML, et al. TNF receptor I sensitizes neurons to erythropoietin- and VEGF-mediated neuroprotection after ischemic and excitotoxic injury. Proc Natl Acad Sci USA. 2008;105:6185–90.
Lu MO, Zhang XM, Mix E, Quezada HC, Jin T, Zhu J, et al. TNF-alpha receptor 1 deficiency enhances kainic acid-induced hippocampal injury in mice. J Neurosci Res. 2008;86:1608–14.
Balosso S, Ravizza T, Perego C, Peschon J, Campbell IL, De Simoni MG, et al. Tumor necrosis factor-alpha inhibits seizures in mice via p75 receptors. Ann Neurol. 2005;57:804–12.
Patel DC, Wallis G, Dahle EJ, McElroy PB, Thomson KE, Tesi RJ, et al. Hippocampal TNFα signaling contributes to seizure generation in an infection-induced mouse model of limbic epilepsy. eNeuro. 2017;4:ENEURO.0105-17.2017.
Marchetti L, Klein M, Schlett K, Pfizenmaier K, Eisel UL. Tumor necrosis factor (TNF)-mediated neuroprotection against glutamate-induced excitotoxicity is enhanced by N-methyl-D-aspartate receptor activation. Essential role of a TNF receptor 2-mediated phosphatidylinositol 3-kinase-dependent NF-kappa B pathway. J Biol Chem. 2004;279:32869–81.
Thompson C, Gary D, Mattson M, Mackenzie A, Robertson GS. Kainic acid-induced naip expression in the hippocampus is blocked in mice lacking TNF receptors. Brain Res Mol Brain Res. 2004;123:126–31.
Zhang XM, Zheng XY, Sharkawi SS, Ruan Y, Amir N, Azimullah S, et al. Possible protecting role of TNF-α in kainic acid-induced neurotoxicity via down-regulation of NFκB signaling pathway. Curr Alzheimer Res. 2013;10:660–9.
Dolga AM, Granic I, Blank T, Knaus HG, Spiess J, Luiten PG, et al. TNF-alpha-mediates neuroprotection against glutamate-induced excitotoxicity via NF-kappaB-dependent up-regulation of K2.2 channels. J Neurochem. 2008;107:1158–67.
Ettcheto M, Junyent F, de Lemos L, Pallas M, Folch J, Beas-Zarate C, et al. Mice lacking functional fas death receptors are protected from kainic acid-induced apoptosis in the hippocampus. Mol Neurobiol. 2015;52:120–9.
Papazian I, Tsoukala E, Boutou A, Karamita M, Kambas K, Iliopoulou L, et al. Fundamentally different roles of neuronal TNF receptors in CNS pathology: TNFR1 and IKKβ promote microglial responses and tissue injury in demyelination while TNFR2 protects against excitotoxicity in mice. J Neuroinflammation. 2021;18:222.
Li T, Lu C, Xia Z, Xiao B, Luo Y. Inhibition of caspase-8 attenuates neuronal death induced by limbic seizures in a cytochrome c-dependent and Smac/DIABLO-independent way. Brain Res. 2006;1098:204–11.
Henshall DC, Bonislawski DP, Skradski SL, Lan JQ, Meller R, Simon RP. Cleavage of bid may amplify caspase-8-induced neuronal death following focally evoked limbic seizures. Neurobiol Dis. 2001;8:568–80.
Lee P, Sata M, Lefer DJ, Factor SM, Walsh K, Kitsis RN. Fas pathway is a critical mediator of cardiac myocyte death and MI during ischemia-reperfusion in vivo. Am J Physiol Heart Circ Physiol. 2003;284:H456–463.
Jeremias I, Kupatt C, Martin-Villalba A, Habazettl H, Schenkel J, Boekstegers P, et al. Involvement of CD95/Apo1/Fas in cell death after myocardial ischemia. Circulation. 2000;102:915–20.
Tekin D, Xi L, Kukreja RC. Genetic deletion of fas receptors or Fas ligands does not reduce infarct size after acute global ischemia-reperfusion in isolated mouse heart. Cell Biochem Biophys. 2006;44:111–7.
Boisguérin P, Covinhes A, Gallot L, Barrère C, Vincent A, Busson M, et al. A novel therapeutic peptide targeting myocardial reperfusion injury. Cardiovasc Res. 2020;116:633–44.
Shiraishi H, Toyozaki T, Tsukamoto Y, Saito T, Masuda Y, Hiroshima K, et al. Antibody binding to fas ligand attenuates inflammatory cell infiltration and cytokine secretion, leading to reduction of myocardial infarct areas and reperfusion injury. Lab Investig. 2002;82:1121–9.
Covinhes A, Gallot L, Barrère C, Vincent A, Sportouch C, Piot C, et al. Anti-apoptotic peptide for long term cardioprotection in a mouse model of myocardial ischemia-reperfusion injury. Sci Rep. 2020;10:18116.
Wang Y, Zhang H, Wang Z, Wei Y, Wang M, Liu M, et al. Blocking the death checkpoint protein TRAIL improves cardiac function after myocardial infarction in monkeys, pigs, and rats. Sci Transl Med. 2020;12:eaaw3172.
Mattisson IY, Björkbacka H, Wigren M, Edsfeldt A, Melander O, Fredrikson GN, et al. Elevated markers of death receptor-activated apoptosis are associated with increased risk for development of diabetes and cardiovascular disease. EBioMedicine. 2017;26:187–97.
Stenemo M, Nowak C, Byberg L, Sundström J, Giedraitis V, Lind L, et al. Circulating proteins as predictors of incident heart failure in the elderly. Eur J Heart Fail. 2018;20:55–62.
Tanner MA, Thomas TP, Grisanti LA. Death receptor 5 contributes to cardiomyocyte hypertrophy through epidermal growth factor receptor transactivation. J Mol Cell Cardiol. 2019;136:1–14.
Di Bartolo BA, Cartland SP, Prado-Lourenco L, Griffith TS, Gentile C, Ravindran J, et al. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) promotes angiogenesis and ischemia-induced neovascularization via NADPH oxidase 4 (NOX4) and nitric oxide-dependent mechanisms. J Am Heart Assoc. 2015;4:e002527.
Hamid T, Gu Y, Ortines RV, Bhattacharya C, Wang G, Xuan YT, et al. Divergent tumor necrosis factor receptor-related remodeling responses in heart failure: role of nuclear factor-kappaB and inflammatory activation. Circulation. 2009;119:1386–97.
Zhang Y, Zhao J, Lau WB, Jiao LY, Liu B, Yuan Y, et al. Tumor necrosis factor-α and lymphotoxin-α mediate myocardial ischemic injury via TNF receptor 1, but are cardioprotective when activating TNF receptor 2. PLoS ONE. 2013;8:e60227.
Kelly ML, Wang M, Crisostomo PR, Abarbanell AM, Herrmann JL, Weil BR, et al. TNF receptor 2, not TNF receptor 1, enhances mesenchymal stem cell-mediated cardiac protection following acute ischemia. Shock. 2010;33:602–7.
Monden Y, Kubota T, Inoue T, Tsutsumi T, Kawano S, Ide T, et al. Tumor necrosis factor-alpha is toxic via receptor 1 and protective via receptor 2 in a murine model of myocardial infarction. Am J Physiol Heart Circ Physiol. 2007;293:H743–753.
Luo D, Luo Y, He Y, Zhang H, Zhang R, Li X, et al. Differential functions of tumor necrosis factor receptor 1 and 2 signaling in ischemia-mediated arteriogenesis and angiogenesis. Am J Pathol. 2006;169:1886–98.
Gouweleeuw L, Wajant H, Maier O, Eisel ULM, Blankesteijn WM, Schoemaker RG. Effects of selective TNFR1 inhibition or TNFR2 stimulation, compared to non-selective TNF inhibition, on (neuro)inflammation and behavior after myocardial infarction in male mice. Brain Behav Immun. 2021;93:156–71.
Guo X, Yin H, Li L, Chen Y, Li J, Doan J, et al. Cardioprotective role of tumor necrosis factor receptor-associated factor 2 by suppressing apoptosis and necroptosis. Circulation. 2017;136:729–42.
Higuchi Y, McTiernan CF, Frye CB, McGowan BS, Chan TO, Feldman AM. Tumor necrosis factor receptors 1 and 2 differentially regulate survival, cardiac dysfunction, and remodeling in transgenic mice with tumor necrosis factor-alpha-induced cardiomyopathy. Circulation. 2004;109:1892–7.
Mann DL, McMurray JJ, Packer M, Swedberg K, Borer JS, Colucci WS, et al. Targeted anticytokine therapy in patients with chronic heart failure: results of the Randomized Etanercept Worldwide Evaluation (RENEWAL). Circulation. 2004;109:1594–602.
Deswal A, Bozkurt B, Seta Y, Parilti-Eiswirth S, Hayes FA, Blosch C, et al. Safety and efficacy of a soluble P75 tumor necrosis factor receptor (Enbrel, etanercept) in patients with advanced heart failure. Circulation. 1999;99:3224–6.
Bozkurt B, Torre-Amione G, Warren MS, Whitmore J, Soran OZ, Feldman AM, et al. Results of targeted anti-tumor necrosis factor therapy with etanercept (ENBREL) in patients with advanced heart failure. Circulation. 2001;103:1044–7.
Generali E, Carrara G, Kallikourdis M, Condorelli G, Bortoluzzi A, Scirè CA, et al. Risk of hospitalization for heart failure in rheumatoid arthritis patients treated with etanercept and abatacept. Rheumatol Int. 2019;39:239–43.
Fan Q, Huang ZM, Boucher M, Shang X, Zuo L, Brinks H, et al. Inhibition of Fas-associated death domain-containing protein (FADD) protects against myocardial ischemia/reperfusion injury in a heart failure mouse model. PLoS ONE. 2013;8:e73537.
Xiao J, Moon M, Yan L, Nian M, Zhang Y, Liu C, et al. Cellular FLICE-inhibitory protein protects against cardiac remodelling after myocardial infarction. Basic Res Cardiol. 2012;107:239.
Liu D, Wu H, Li YZ, Yang J, Yang J, Ding JW, et al. Cellular FADD-like IL-1β-converting enzyme-inhibitory protein attenuates myocardial ischemia/reperfusion injury via suppressing apoptosis and autophagy simultaneously. Nutr Metab Cardiovasc Dis. 2021;31:1916–28.
Liang Y, Lin Q, Zhu J, Li X, Fu Y, Zou X, et al. The caspase-8 shRNA-modified mesenchymal stem cells improve the function of infarcted heart. Mol Cell Biochem. 2014;397:7–16.
Fauconnier J, Meli AC, Thireau J, Roberge S, Shan J, Sassi Y, et al. Ryanodine receptor leak mediated by caspase-8 activation leads to left ventricular injury after myocardial ischemia-reperfusion. Proc Natl Acad Sci USA. 2011;108:13258–63.
Scharner D, Rössig L, Carmona G, Chavakis E, Urbich C, Fischer A, et al. Caspase-8 is involved in neovascularization-promoting progenitor cell functions. Arterioscler Thromb Vasc Biol. 2009;29:571–8.
Koshinuma S, Miyamae M, Kaneda K, Kotani J, Figueredo VM. Combination of necroptosis and apoptosis inhibition enhances cardioprotection against myocardial ischemia-reperfusion injury. J Anesth. 2014;28:235–41.
Nam SW, Liu H, Wong JZ, Feng AY, Chu G, Merchant N, et al. Cardiomyocyte apoptosis contributes to pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated mice. Clin Sci. 2014;127:519–26.
Toffoli B, Bernardi S, Candido R, Zacchigna S, Fabris B, Secchiero P. TRAIL shows potential cardioprotective activity. Investig N Drugs. 2012;30:1257–60.
Papathanasiou S, Rickelt S, Soriano ME, Schips TG, Maier HJ, Davos CH, et al. Tumor necrosis factor-α confers cardioprotection through ectopic expression of keratins K8 and K18. Nat Med. 2015;21:1076–84.
Liu Z, Fitzgerald M, Meisinger T, Batra R, Suh M, Greene H, et al. CD95-ligand contributes to abdominal aortic aneurysm progression by modulating inflammation. Cardiovasc Res. 2019;115:807–18.
Wencker D, Chandra M, Nguyen K, Miao W, Garantziotis S, Factor SM, et al. A mechanistic role for cardiac myocyte apoptosis in heart failure. J Clin Investig. 2003;111:1497–504.
Tanner MA, Grisanti LA. A dual role for death receptor 5 in regulating cardiac fibroblast function. Front Cardiovasc Med. 2021;8:699102.
Pircher J, Merkle M, Wörnle M, Ribeiro A, Czermak T, Stampnik Y, et al. Prothrombotic effects of tumor necrosis factor alpha in vivo are amplified by the absence of TNF-alpha receptor subtype 1 and require TNF-alpha receptor subtype 2. Arthritis Res Ther. 2012;14:R225.
Duerrschmid C, Crawford JR, Reineke E, Taffet GE, Trial J, Entman ML, et al. TNF receptor 1 signaling is critically involved in mediating angiotensin-II-induced cardiac fibrosis. J Mol Cell Cardiol. 2013;57:59–67.
Woods C, Marques-Lopes J, Contoreggi NH, Milner TA, Pickel VM, Wang G, et al. Tumor necrosis factor α receptor type 1 activation in the hypothalamic paraventricular nucleus contributes to glutamate signaling and angiotensin II-dependent hypertension. J Neurosci. 2021;41:1349–62.
Li H, Tang QZ, Liu C, Moon M, Chen M, Yan L, et al. Cellular FLICE-inhibitory protein protects against cardiac remodeling induced by angiotensin II in mice. Hypertension. 2010;56:1109–17.
Jobe LJ, Meléndez GC, Levick SP, Du Y, Brower GL, Janicki JS. TNF-alpha inhibition attenuates adverse myocardial remodeling in a rat model of volume overload. Am J Physiol Heart Circ Physiol. 2009;297:H1462–1468.
Sun M, Chen M, Dawood F, Zurawska U, Li JY, Parker T, et al. Tumor necrosis factor-alpha mediates cardiac remodeling and ventricular dysfunction after pressure overload state. Circulation. 2007;115:1398–407.
Badorff C, Ruetten H, Mueller S, Stahmer M, Gehring D, Jung F, et al. Fas receptor signaling inhibits glycogen synthase kinase 3 beta and induces cardiac hypertrophy following pressure overload. J Clin Investig. 2002;109:373–81.
Stamm C, Friehs I, Cowan DB, Moran AM, Cao-Danh H, Duebener LF, et al. Inhibition of tumor necrosis factor-alpha improves postischemic recovery of hypertrophied hearts. Circulation. 2001;104:I350–355.
Miao K, Zhou L, Ba H, Li C, Gu H, Yin B, et al. Transmembrane tumor necrosis factor alpha attenuates pressure-overload cardiac hypertrophy via tumor necrosis factor receptor 2. PLoS Biol. 2020;18:e3000967.
Mattos BR, Bonacio GF, Vitorino TR, Garcia VT, Amaral JH, Dellalibera-Joviliano R, et al. TNF-α inhibition decreases MMP-2 activity, reactive oxygen species formation and improves hypertensive vascular hypertrophy independent of its effects on blood pressure. Biochem Pharmacol. 2020;180:114121.
Giampietri C, Petrungaro S, Musumeci M, Coluccia P, Antonangeli F, De Cesaris P, et al. c-Flip overexpression reduces cardiac hypertrophy in response to pressure overload. J Hypertens. 2008;26:1008–16.
Hsu CC, Li Y, Hsu CT, Cheng JT, Lin MH, Cheng KC, et al. Etanercept ameliorates cardiac fibrosis in rats with diet-induced obesity. Pharmaceuticals. 2021;14:320.
Di Bartolo BA, Cartland SP, Harith HH, Bobryshev YV, Schoppet M, Kavurma MM. TRAIL-deficiency accelerates vascular calcification in atherosclerosis via modulation of RANKL. PLoS ONE. 2013;8:e74211.
Di Bartolo BA, Chan J, Bennett MR, Cartland S, Bao S, Tuch BE, et al. TNF-related apoptosis-inducing ligand (TRAIL) protects against diabetes and atherosclerosis in Apoe −/− mice. Diabetologia. 2011;54:3157–67.
Watt V, Chamberlain J, Steiner T, Francis S, Crossman D. TRAIL attenuates the development of atherosclerosis in apolipoprotein E deficient mice. Atherosclerosis. 2011;215:348–54.
Zadelaar AS, von der Thüsen JH, Boesten LS, Hoeben RC, Kockx MM, Versnel MA, et al. Increased vulnerability of pre-existing atherosclerosis in ApoE-deficient mice following adenovirus-mediated Fas ligand gene transfer. Atherosclerosis. 2005;183:244–50.
Yang J, Sato K, Aprahamian T, Brown NJ, Hutcheson J, Bialik A, et al. Endothelial overexpression of Fas ligand decreases atherosclerosis in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 2004;24:1466–73.
Xanthoulea S, Thelen M, Pöttgens C, Gijbels MJ, Lutgens E, de Winther MP. Absence of p55 TNF receptor reduces atherosclerosis, but has no major effect on angiotensin II induced aneurysms in LDL receptor deficient mice. PLoS ONE. 2009;4:e6113.
Xanthoulea S, Gijbels MJ, van der Made I, Mujcic H, Thelen M, Vergouwe MN, et al. P55 tumour necrosis factor receptor in bone marrow-derived cells promotes atherosclerosis development in low-density lipoprotein receptor knock-out mice. Cardiovasc Res. 2008;80:309–18.
Zhang L, Peppel K, Sivashanmugam P, Orman ES, Brian L, Exum ST, et al. Expression of tumor necrosis factor receptor-1 in arterial wall cells promotes atherosclerosis. Arterioscler Thromb Vasc Biol. 2007;27:1087–94.
Brånén L, Hovgaard L, Nitulescu M, Bengtsson E, Nilsson J, Jovinge S. Inhibition of tumor necrosis factor-alpha reduces atherosclerosis in apolipoprotein E knockout mice. Arterioscler Thrombos Vasc Biol. 2004;24:2137–42.
Miyata S, Takemura G, Kosai K, Takahashi T, Esaki M, Li L, et al. Anti-Fas gene therapy prevents doxorubicin-induced acute cardiotoxicity through mechanisms independent of apoptosis. Am J Pathol. 2010;176:687–98.
Niu J, Azfer A, Wang K, Wang X, Kolattukudy PE. Cardiac-targeted expression of soluble fas attenuates doxorubicin-induced cardiotoxicity in mice. J Pharmacol Exp Ther. 2009;328:740–8.
Clayton ZS, Brunt VE, Hutton DA, Casso AG, Ziemba BP, Melov S, et al. Tumor necrosis factor alpha-mediated inflammation and remodeling of the extracellular matrix underlies aortic stiffening induced by the common chemotherapeutic agent doxorubicin. Hypertension. 2021;77:1581–90.
Furuichi K, Kokubo S, Hara A, Imamura R, Wang Q, Kitajima S, et al. Fas ligand has a greater impact than TNF-α on apoptosis and inflammation in ischemic acute kidney injury. Nephron Extra. 2012;2:27–38.
Ko GJ, Jang HR, Huang Y, Womer KL, Liu M, Higbee E, et al. Blocking Fas ligand on leukocytes attenuates kidney ischemia-reperfusion injury. J Am Soc Nephrology. 2011;22:732–42.
Hamar P, Song E, Kökény G, Chen A, Ouyang N, Lieberman J. Small interfering RNA targeting Fas protects mice against renal ischemia-reperfusion injury. Proc Natl Acad Sci USA. 2004;101:14883–8.
Du C, Wang S, Diao H, Guan Q, Zhong R, Jevnikar AM. Increasing resistance of tubular epithelial cells to apoptosis by shRNA therapy ameliorates renal ischemia-reperfusion injury. Am J Transplant. 2006;6:2256–67.
Hou L, Chen G, Feng B, Zhang XS, Zheng XF, Xiang Y, et al. Small interfering RNA targeting TNF-α gene significantly attenuates renal ischemia-reperfusion injury in mice. J Huazhong Univ Sci Technol Med Sci. 2016;36:634–8.
Adachi T, Sugiyama N, Yagita H, Yokoyama T. Renal atrophy after ischemia-reperfusion injury depends on massive tubular apoptosis induced by TNFα in the later phase. Med Mol Morphol. 2014;47:213–23.
Choi DE, Jeong JY, Lim BJ, Na KR, Shin YT, Lee KW. Pretreatment with the tumor nerosis factor-alpha blocker etanercept attenuated ischemia-reperfusion renal injury. Transplant Proc. 2009;41:3590–6.
Adachi T, Sugiyama N, Gondai T, Yagita H, Yokoyama T. Blockade of Death Ligand TRAIL Inhibits Renal Ischemia Reperfusion Injury. Acta Histochem Cytochem. 2013;46:161–70.
Leng X, Zhang Q, Chen Z, Wang D. Blocking TRAIL-DR5 signaling with soluble DR5 alleviates acute kidney injury in a severely burned mouse model. Int J Clin Exp Pathol. 2014;7:3460–8.
Ramesh G, Reeves WB. TNF-alpha mediates chemokine and cytokine expression and renal injury in cisplatin nephrotoxicity. J Clin Investig. 2002;110:835–42.
Lin JS, Mamlouk O, Selamet U, Tchakarov A, Glass WF, Sheth RA, et al. Infliximab for the treatment of patients with checkpoint inhibitor-associated acute tubular interstitial nephritis. Oncoimmunology. 2021;10:1877415.
Misaki T, Yamamoto T, Suzuki S, Fukasawa H, Togawa A, Ohashi N, et al. Decrease in tumor necrosis factor-alpha receptor-associated death domain results from ubiquitin-dependent degradation in obstructive renal injury in rats. Am J Pathol. 2009;175:74–83.
Misseri R, Meldrum DR, Dinarello CA, Dagher P, Hile KL, Rink RC, et al. TNF-alpha mediates obstruction-induced renal tubular cell apoptosis and proapoptotic signaling. Am J Physiol Ren Physiol. 2005;288:F406–411.
Morimoto Y, Gai Z, Tanishima H, Kawakatsu M, Itoh S, Hatamura I, et al. TNF-alpha deficiency accelerates renal tubular interstitial fibrosis in the late stage of ureteral obstruction. Exp Mol Pathol. 2008;85:207–13.
Guo G, Morrissey J, McCracken R, Tolley T, Klahr S. Role of TNFR1 and TNFR2 receptors in tubulointerstitial fibrosis of obstructive nephropathy. Am J Physiol. 1999;277:F766–772.
Hughes J, Johnson RJ. Role of Fas (CD95) in tubulointerstitial disease induced by unilateral ureteric obstruction. Am J Physiol. 1999;277:F26–32.
Zhang X, Zheng X, Sun H, Feng B, Chen G, Vladau C, et al. Prevention of renal ischemic injury by silencing the expression of renal caspase 3 and caspase 8. Transplantation. 2006;82:1728–32.
Linkermann A, Bräsen JH, Himmerkus N, Liu S, Huber TB, Kunzendorf U, et al. Rip1 (receptor-interacting protein kinase 1) mediates necroptosis and contributes to renal ischemia/reperfusion injury. Kidney Int. 2012;81:751–61.
Linkermann A, Bräsen JH, Darding M, Jin MK, Sanz AB, Heller JO, et al. Two independent pathways of regulated necrosis mediate ischemia-reperfusion injury. Proc Natl Acad Sci USA. 2013;110:12024–9.
Sung B, Su Y, Jiang J, McLeod P, Liu W, Haig A, et al. Loss of receptor interacting protein kinases 3 and caspase-8 augments intrinsic apoptosis in tubular epithelial cell and promote kidney ischaemia-reperfusion injury. Nephrol. 2019;24:661–9.
Awad AS, You H, Gao T, Cooper TK, Nedospasov SA, Vacher J, et al. Macrophage-derived tumor necrosis factor-α mediates diabetic renal injury. Kidney Int. 2015;88:722–33.
Omote K, Gohda T, Murakoshi M, Sasaki Y, Kazuno S, Fujimura T, et al. Role of the TNF pathway in the progression of diabetic nephropathy in KK-A(y) mice. Am J Physiol Ren Physiol. 2014;306:F1335–1347.
Moriwaki Y, Inokuchi T, Yamamoto A, Ka T, Tsutsumi Z, Takahashi S, et al. Effect of TNF-alpha inhibition on urinary albumin excretion in experimental diabetic rats. Acta Diabetol. 2007;44:215–8.
Cheng D, Liang R, Huang B, Hou J, Yin J, Zhao T, et al. Tumor necrosis factor-α blockade ameliorates diabetic nephropathy in rats. Clin Kidney J. 2021;14:301–8.
Cartland SP, Erlich JH, Kavurma MM. TRAIL deficiency contributes to diabetic nephropathy in fat-fed ApoE−/− mice. PLoS ONE. 2014;9:e92952.
Lorz C, Benito-Martín A, Boucherot A, Ucero AC, Rastaldi MP, Henger A, et al. The death ligand TRAIL in diabetic nephropathy. J Am Soc Nephrology. 2008;19:904–14.
Toffoli B, Tonon F, Tisato V, Michelli A, Zauli G, Secchiero P, et al. TRAIL treatment prevents renal morphological changes and TGF-β-induced mesenchymal transition associated with diabetic nephropathy. Clin Sci. 2020;134:2337–52.
Roix J, Saha S. TNF-α blockade is ineffective in animal models of established polycystic kidney disease. BMC Nephrol. 2013;14:233.
Li X, Magenheimer BS, Xia S, Johnson T, Wallace DP, Calvet JP, et al. A tumor necrosis factor-alpha-mediated pathway promoting autosomal dominant polycystic kidney disease. Nat Med. 2008;14:863–8.
Tarzi RM, Sharp PE, McDaid JP, Fossati-Jimack L, Herbert PE, Pusey CD, et al. Mice with defective Fas ligand are protected from crescentic glomerulonephritis. Kidney Int. 2012;81:170–8.
Khan SB, Cook HT, Bhangal G, Smith J, Tam FW, Pusey CD. Antibody blockade of TNF-alpha reduces inflammation and scarring in experimental crescentic glomerulonephritis. Kidney Int. 2005;67:1812–20.
Zaenker M, Arbach O, Helmchen U, Glorius P, Ludewig S, Braasch E. Crescentic glomerulonephritis associated with myeloperoxidase-antineutrophil-cytoplasmic antibodies: first report on the efficacy of primary anti-TNF-alpha treatment. Int J tissue React. 2004;26:85–92.
Le Hir M, Haas C, Marino M, Ryffel B. Prevention of crescentic glomerulonephritis induced by anti-glomerular membrane antibody in tumor necrosis factor-deficient mice. Lab Investig. 1998;78:1625–31.
Wen Y, Rudemiller NP, Zhang J, Robinette T, Lu X, Ren J, et al. TNF-α in T lymphocytes attenuates renal injury and fibrosis during nephrotoxic nephritis. Am J Physiol Ren Physiol. 2020;318:F107–f116.
Taubitz A, Schwarz M, Eltrich N, Lindenmeyer MT, Vielhauer V. Distinct contributions of TNF receptor 1 and 2 to TNF-induced glomerular inflammation in mice. PLoS ONE. 2013;8:e68167.
Pfeifer E, Polz J, Grieβl S, Mostböck S, Hehlgans T, Männel DN. Mechanisms of immune complex-mediated experimental glomerulonephritis: possible role of the balance between endogenous TNF and soluble TNF receptor type 2. Eur Cytokine Netw. 2012;23:15–20.
Vielhauer V, Stavrakis G, Mayadas TN. Renal cell-expressed TNF receptor 2, not receptor 1, is essential for the development of glomerulonephritis. J Clin Investig. 2005;115:1199–209.
Ryffel B, Eugster H, Haas C, Le, Hir M. Failure to induce anti-glomerular basement membrane glomerulonephritis in TNF alpha/beta deficient mice. Int J Exp Pathol. 1998;79:453–60.
Müller MB, Hoppe JM, Bideak A, Lux M, Lindenmeyer MT, Müller S, et al. Exclusive expression of transmembrane TNF aggravates acute glomerulonephritis despite reduced leukocyte infiltration and inflammation. Kidney Int. 2019;95:75–93.
Mahmoud MF, El Shazly SM, Barakat W. Inhibition of TNF-α protects against hepatic ischemia-reperfusion injury in rats via NF-κB dependent pathway. Naunyn Schmiedebergs Arch Pharmacol. 2012;385:465–71.
Hernandez-Alejandro R, Zhang X, Croome KP, Zheng X, Parfitt J, Chen D, et al. Reduction of liver ischemia reperfusion injury by silencing of TNF-α gene with shRNA. J Surg Res. 2012;176:614–20.
Rüdiger HA, Clavien PA. Tumor necrosis factor alpha, but not Fas, mediates hepatocellular apoptosis in the murine ischemic liver. Gastroenterology. 2002;122:202–10.
Al-Saeedi M, Steinebrunner N, Kudsi H, Halama N, Mogler C, Büchler MW, et al. Neutralization of CD95 ligand protects the liver against ischemia-reperfusion injury and prevents acute liver failure. Cell Death Dis. 2018;9:132.
Nakajima H, Mizuta N, Fujiwara I, Sakaguchi K, Ogata H, Magae J, et al. Blockade of the Fas/Fas ligand interaction suppresses hepatocyte apoptosis in ischemia-reperfusion rat liver. Apoptosis. 2008;13:1013–21.
Teoh N, Leclercq I, Pena AD, Farrell G. Low-dose TNF-alpha protects against hepatic ischemia-reperfusion injury in mice: implications for preconditioning. Hepatology. 2003;37:118–28.
Fahrner R, Trochsler M, Corazza N, Graubardt N, Keogh A, Candinas D, et al. Tumor necrosis factor-related apoptosis-inducing ligand on NK cells protects from hepatic ischemia-reperfusion injury. Transplantation. 2014;97:1102–9.
Contreras JL, Vilatoba M, Eckstein C, Bilbao G, Anthony Thompson J, Eckhoff DE. Caspase-8 and caspase-3 small interfering RNA decreases ischemia/reperfusion injury to the liver in mice. Surgery. 2004;136:390–400.
Kolachala VL, Palle SK, Shen M, Shenoi A, Shayakhmetov DM, Gupta NA. Influence of fat on differential receptor interacting serine/threonine protein kinase 1 activity leading to apoptotic cell death in murine liver ischemia reperfusion injury through caspase 8. Hepatol Commun. 2019;3:925–42.
Williams CD, McGill MR, Farhood A, Jaeschke H. Fas receptor-deficient lpr mice are protected against acetaminophen hepatotoxicity due to higher glutathione synthesis and enhanced detoxification of oxidant stress. Food Chem Toxicol. 2013;58:228–35.
Chen Q, Yan D, Zhang Q, Zhang G, Xia M, Li J, et al. Treatment of acetaminophen-induced liver failure by blocking the death checkpoint protein TRAIL. Biochim Biophys Acta Mol Basis Dis. 2020;1866:165583.
Tinel M, Berson A, Vadrot N, Descatoire V, Grodet A, Feldmann G, et al. Subliminal Fas stimulation increases the hepatotoxicity of acetaminophen and bromobenzene in mice. Hepatology. 2004;39:655–66.
Schattenberg JM, Nagel M, Kim YO, Kohl T, Wörns MA, Zimmermann T, et al. Increased hepatic fibrosis and JNK2-dependent liver injury in mice exhibiting hepatocyte-specific deletion of cFLIP. Am J Physiol Gastrointest Liver Physiol. 2012;303:G498–506.
Schuchmann M, Varfolomeev EE, Hermann F, Rueckert F, Strand D, Koehler H, et al. Dominant negative MORT1/FADD rescues mice from CD95 and TNF-induced liver failure. Hepatology. 2003;37:129–35.
Seino K, Setoguchi Y, Ogino T, Kayagaki N, Akiba H, Nakano H, et al. Protection against Fas-mediated and tumor necrosis factor receptor 1-mediated liver injury by blockade of FADD without loss of nuclear factor-kappaB activation. Ann Surg. 2001;234:681–8.
Liedtke C, Bangen JM, Freimuth J, Beraza N, Lambertz D, Cubero FJ, et al. Loss of caspase-8 protects mice against inflammation-related hepatocarcinogenesis but induces non-apoptotic liver injury. Gastroenterology. 2011;141:2176–87.
Ni HM, McGill MR, Chao X, Woolbright BL, Jaeschke H, Ding WX. Caspase inhibition prevents tumor necrosis factor-α-induced apoptosis and promotes necrotic cell death in mouse hepatocytes in vivo and in vitro. Am J Pathol. 2016;186:2623–36.
Wroblewski R, Armaka M, Kondylis V, Pasparakis M, Walczak H, Mittrücker HW, et al. Opposing role of tumor necrosis factor receptor 1 signaling in T cell-mediated hepatitis and bacterial infection in mice. Hepatology. 2016;64:508–21.
Zender L, Hutker S, Liedtke C, Tillmann HL, Zender S, Mundt B, et al. Caspase 8 small interfering RNA prevents acute liver failure in mice. Proc Natl Acad Sci USA. 2003;100:7797–802.
Schattenberg JM, Zimmermann T, Wörns M, Sprinzl MF, Kreft A, Kohl T, et al. Ablation of c-FLIP in hepatocytes enhances death-receptor mediated apoptosis and toxic liver injury in vivo. J Hepatol. 2011;55:1272–80.
Lazic M, Eguchi A, Berk MP, Povero D, Papouchado B, Mulya A, et al. Differential regulation of inflammation and apoptosis in Fas-resistant hepatocyte-specific Bid-deficient mice. J Hepatol. 2014;61:107–15.
Kaufmann T, Tai L, Ekert PG, Huang DC, Norris F, Lindemann RK, et al. The BH3-only protein bid is dispensable for DNA damage- and replicative stress-induced apoptosis or cell-cycle arrest. Cell. 2007;129:423–33.
Hikita H, Takehara T, Kodama T, Shimizu S, Shigekawa M, Hosui A, et al. Delayed-onset caspase-dependent massive hepatocyte apoptosis upon Fas activation in Bak/Bax-deficient mice. Hepatology. 2011;54:240–51.
Rodriguez I, Matsuura K, Khatib K, Reed JC, Nagata S, Vassalli P. A bcl-2 transgene expressed in hepatocytes protects mice from fulminant liver destruction but not from rapid death induced by anti-Fas antibody injection. J Exp Med. 1996;183:1031–6.
Lacronique V, Mignon A, Fabre M, Viollet B, Rouquet N, Molina T, et al. Bcl-2 protects from lethal hepatic apoptosis induced by an anti-Fas antibody in mice. Nat Med. 1996;2:80–86.
Tan S, Liu X, Chen L, Wu X, Tao L, Pan X, et al. Fas/FasL mediates NF-κBp65/PUMA-modulated hepatocytes apoptosis via autophagy to drive liver fibrosis. Cell Death Dis. 2021;12:474.
Yan J, Xiang J, Lin Y, Ma J, Zhang J, Zhang H, et al. Inactivation of BAD by IKK inhibits TNFα-induced apoptosis independently of NF-κB activation. Cell. 2013;152:304–15.
Ottina E, Sochalska M, Sgonc R, Villunger A. The BH3-only protein Bad is dispensable for TNF-mediated cell death. Cell Death Dis. 2015;6:e1611.
Woo M, Hakem A, Elia AJ, Hakem R, Duncan GS, Patterson BJ, et al. In vivo evidence that caspase-3 is required for Fas-mediated apoptosis of hepatocytes. J Immunol. 1999;163:4909–16.
Bajt ML, Vonderfecht SL, Jaeschke H. Differential protection with inhibitors of caspase-8 and caspase-3 in murine models of tumor necrosis factor and Fas receptor-mediated hepatocellular apoptosis. Toxicol Appl Pharmacol. 2001;175:243–52.
Zheng TS, Hunot S, Kuida K, Momoi T, Srinivasan A, Nicholson DW, et al. Deficiency in caspase-9 or caspase-3 induces compensatory caspase activation. Nat Med. 2000;6:1241–7.
Sudo K, Yamada Y, Saito K, Shimizu S, Ohashi H, Kato T, et al. TNF-alpha and IL-6 signals from the bone marrow derived cells are necessary for normal murine liver regeneration. Biochim Biophys Acta. 2008;1782:671–9.
Desbarats J, Newell MK. Fas engagement accelerates liver regeneration after partial hepatectomy. Nat Med. 2000;6:920–3.
Knight B, Yeoh GC. TNF/LTalpha double knockout mice display abnormal inflammatory and regenerative responses to acute and chronic liver injury. Cell Tissue Res. 2005;319:61–70.
Taira K, Hiroyasu S, Shiraishi M, Muto Y, Koji T. Role of the Fas system in liver regeneration after a partial hepatectomy in rats. Eur Surg Res. 2001;33:334–41.
Schuchmann M, Ruckert F, Garcia-Lazaro JF, Karg A, Burg J, Knorr N, et al. MORT1/FADD is involved in liver regeneration. World J Gastroenterol. 2005;11:7248–53.
Ben Moshe T, Barash H, Kang TB, Kim JC, Kovalenko A, Gross E, et al. Role of caspase-8 in hepatocyte response to infection and injury in mice. Hepatology. 2007;45:1014–24.
Freimuth J, Bangen JM, Lambertz D, Hu W, Nevzorova YA, Sonntag R, et al. Loss of caspase-8 in hepatocytes accelerates the onset of liver regeneration in mice through premature nuclear factor kappa B activation. Hepatology. 2013;58:1779–89.
Isayama F, Moore S, Hines IN, Wheeler MD. Fas regulates macrophage polarization and fibrogenic phenotype in a model of chronic ethanol-induced hepatocellular injury. Am J Pathol. 2016;186:1524–36.
Verma VK, Li H, Wang R, Hirsova P, Mushref M, Liu Y, et al. Alcohol stimulates macrophage activation through caspase-dependent hepatocyte derived release of CD40L containing extracellular vesicles. J Hepatol. 2016;64:651–60.
Yin M, Wheeler MD, Kono H, Bradford BU, Gallucci RM, Luster MI, et al. Essential role of tumor necrosis factor alpha in alcohol-induced liver injury in mice. Gastroenterology. 1999;117:942–52.
Mundt B, Wirth T, Zender L, Waltemathe M, Trautwein C, Manns MP, et al. Tumour necrosis factor related apoptosis inducing ligand (TRAIL) induces hepatic steatosis in viral hepatitis and after alcohol intake. Gut. 2005;54:1590–6.
Hao F, Cubero FJ, Ramadori P, Liao L, Haas U, Lambertz D, et al. Inhibition of Caspase-8 does not protect from alcohol-induced liver apoptosis but alleviates alcoholic hepatic steatosis in mice. Cell Death Dis. 2017;8:e3152.
Zhou Z, Sun X, Kang YJ. Ethanol-induced apoptosis in mouse liver: Fas- and cytochrome c-mediated caspase-3 activation pathway. Am J Pathol. 2001;159:329–38.
Item F, Wueest S, Lemos V, Stein S, Lucchini FC, Denzler R, et al. Fas cell surface death receptor controls hepatic lipid metabolism by regulating mitochondrial function. Nat Commun. 2017;8:480.
Kakino S, Ohki T, Nakayama H, Yuan X, Otabe S, Hashinaga T, et al. Pivotal role of TNF-α in the development and progression of nonalcoholic fatty liver disease in a murine model. Horm Metab Res. 2018;50:80–87.
Salles J, Tardif N, Landrier JF, Mothe-Satney I, Guillet C, Boue-Vaysse C, et al. TNFα gene knockout differentially affects lipid deposition in liver and skeletal muscle of high-fat-diet mice. J Nutr Biochem. 2012;23:1685–93.
Kanuri G, Spruss A, Wagnerberger S, Bischoff SC, Bergheim I. Role of tumor necrosis factor α (TNFα) in the onset of fructose-induced nonalcoholic fatty liver disease in mice. J Nutr Biochem. 2011;22:527–34.
Tomita K, Tamiya G, Ando S, Ohsumi K, Chiyo T, Mizutani A, et al. Tumour necrosis factor alpha signalling through activation of Kupffer cells plays an essential role in liver fibrosis of non-alcoholic steatohepatitis in mice. Gut. 2006;55:415–24.
De Sousa Rodrigues ME, Houser MC, Walker DI, Jones DP, Chang J, Barnum CJ, et al. Targeting soluble tumor necrosis factor as a potential intervention to lower risk for late-onset Alzheimer’s disease associated with obesity, metabolic syndrome, and type 2 diabetes. Alzheimers Res Ther. 2019;12:1.
Ilan Y, Ben Ya’acovA, Shabbat Y, Gingis-Velitski S, Almon E, Shaaltiel Y. Oral administration of a non-absorbable plant cell-expressed recombinant anti-TNF fusion protein induces immunomodulatory effects and alleviates nonalcoholic steatohepatitis. World J Gastroenterol. 2016;22:8760–9.
Koca SS, Bahcecioglu IH, Poyrazoglu OK, Ozercan IH, Sahin K, Ustundag B. The treatment with antibody of TNF-alpha reduces the inflammation, necrosis and fibrosis in the non-alcoholic steatohepatitis induced by methionine- and choline-deficient diet. Inflammation. 2008;31:91–98.
Wandrer F, Liebig S, Marhenke S, Vogel A, John K, Manns MP, et al. TNF-Receptor-1 inhibition reduces liver steatosis, hepatocellular injury and fibrosis in NAFLD mice. Cell Death Dis. 2020;11:212.
Bluemel S, Wang Y, Lee S, Schnabl B. Tumor necrosis factor alpha receptor 1 deficiency in hepatocytes does not protect from non-alcoholic steatohepatitis, but attenuates insulin resistance in mice. World J Gastroenterol. 2020;26:4933–44.
Lambertucci F, Arboatti A, Sedlmeier MG, Motiño O, Alvarez ML, Ceballos MP, et al. Disruption of tumor necrosis factor alpha receptor 1 signaling accelerates NAFLD progression in mice upon a high-fat diet. J Nutr Biochem. 2018;58:17–27.
Bernardi S, Toffoli B, Tisato V, Bossi F, Biffi S, Lorenzon A, et al. TRAIL reduces impaired glucose tolerance and NAFLD in the high-fat diet fed mouse. Clin Sci. 2018;132:69–83.
Hirsova P, Weng P, Salim W, Bronk SF, Griffith TS, Ibrahim SH, et al. TRAIL deletion prevents liver, but not adipose tissue, inflammation during murine diet-induced obesity. Hepatol Commun. 2017;1:648–62.
Cartland SP, Harith HH, Genner SW, Dang L, Cogger VC, Vellozzi M, et al. Non-alcoholic fatty liver disease, vascular inflammation and insulin resistance are exacerbated by TRAIL deletion in mice. Sci Rep. 2017;7:1898.
Krishnan A, Katsumi T, Guicciardi ME, Azad AI, Ozturk NB, Trussoni CE, et al. Tumor necrosis factor-related apoptosis-inducing ligand receptor deficiency promotes the ductular reaction, macrophage accumulation, and hepatic fibrosis in the Abcb4(−/−) mouse. Am J Pathol. 2020;190:1284–97.
Gujral JS, Liu J, Farhood A, Jaeschke H. Reduced oncotic necrosis in Fas receptor-deficient C57BL/6J-lpr mice after bile duct ligation. Hepatology. 2004;40:998–1007.
Canbay A, Higuchi H, Bronk SF, Taniai M, Sebo TJ, Gores GJ. Fas enhances fibrogenesis in the bile duct ligated mouse: a link between apoptosis and fibrosis. Gastroenterology. 2002;123:1323–30.
Miyoshi H, Rust C, Roberts PJ, Burgart LJ, Gores GJ. Hepatocyte apoptosis after bile duct ligation in the mouse involves Fas. Gastroenterology. 1999;117:669–77.
Osawa Y, Hoshi M, Yasuda I, Saibara T, Moriwaki H, Kozawa O. Tumor necrosis factor-α promotes cholestasis-induced liver fibrosis in the mouse through tissue inhibitor of metalloproteinase-1 production in hepatic stellate cells. PloS one. 2013;8:e65251.
Gäbele E, Froh M, Arteel GE, Uesugi T, Hellerbrand C, Schölmerich J, et al. TNFalpha is required for cholestasis-induced liver fibrosis in the mouse. Biochem Biophys Res Commun. 2009;378:348–53.
Takeda K, Kojima Y, Ikejima K, Harada K, Yamashina S, Okumura K, et al. Death receptor 5 mediated-apoptosis contributes to cholestatic liver disease. Proc Natl Acad Sci USA. 2008;105:10895–10900.
Kahraman A, Barreyro FJ, Bronk SF, Werneburg NW, Mott JL, Akazawa Y, et al. TRAIL mediates liver injury by the innate immune system in the bile duct-ligated mouse. Hepatology. 2008;47:1317–30.
Zhuang H, Wang X, Zha D, Gan Z, Cai F, Du P, et al. FADD is a key regulator of lipid metabolism. EMBO Mol Med. 2016;8:895–918.
Wang PX, Ji YX, Zhang XJ, Zhao LP, Yan ZZ, Zhang P, et al. Targeting CASP8 and FADD-like apoptosis regulator ameliorates nonalcoholic steatohepatitis in mice and nonhuman primates. Nat Med. 2017;23:439–49.
Gehrke N, Nagel M, Straub BK, Wörns MA, Schuchmann M, Galle PR, et al. Loss of cellular FLICE-inhibitory protein promotes acute cholestatic liver injury and inflammation from bile duct ligation. Am J Physiol Gastrointest Liver Physiol. 2018;314:G319–g333.
Chaudhary K, Liedtke C, Wertenbruch S, Trautwein C, Streetz KL. Caspase 8 differentially controls hepatocytes and non-parenchymal liver cells during chronic cholestatic liver injury in mice. J Hepatol. 2013;59:1292–8.
Hatting M, Zhao G, Schumacher F, Sellge G, Al Masaoudi M, Gaβler N, et al. Hepatocyte caspase-8 is an essential modulator of steatohepatitis in rodents. Hepatology. 2013;57:2189–201.
Cubero FJ, Peng J, Liao L, Su H, Zhao G, Zoubek ME, et al. Inactivation of caspase 8 in liver parenchymal cells confers protection against murine obstructive cholestasis. J Hepatol. 2018;69:1326–34.
Kondylis V, Polykratis A, Ehlken H, Ochoa-Callejero L, Straub BK, Krishna-Subramanian S, et al. NEMO prevents steatohepatitis and hepatocellular carcinoma by inhibiting RIPK1 kinase activity-mediated hepatocyte apoptosis. Cancer Cell. 2015;28:582–98.
Ehlken H, Krishna-Subramanian S, Ochoa-Callejero L, Kondylis V, Nadi NE, Straub BK, et al. Death receptor-independent FADD signalling triggers hepatitis and hepatocellular carcinoma in mice with liver parenchymal cell-specific NEMO knockout. Cell Death Differ. 2014;21:1721–32.
Vogel A, Aslan JE, Willenbring H, Klein C, Finegold M, Mount H, et al. Sustained phosphorylation of Bid is a marker for resistance to Fas-induced apoptosis during chronic liver diseases. Gastroenterology. 2006;130:104–19.
Straus SE, Jaffe ES, Puck JM, Dale JK, Elkon KB, Rösen-Wolff A, et al. The development of lymphomas in families with autoimmune lymphoproliferative syndrome with germline Fas mutations and defective lymphocyte apoptosis. Blood. 2001;98:194–200.
Davidson WF, Giese T, Fredrickson TN. Spontaneous development of plasmacytoid tumors in mice with defective Fas-Fas ligand interactions. J Exp Med. 1998;187:1825–38.
Finnberg N, Klein-Szanto AJ, El-Deiry WS. TRAIL-R deficiency in mice promotes susceptibility to chronic inflammation and tumorigenesis. J Clin Investig. 2008;118:111–23.
Zerafa N, Westwood JA, Cretney E, Mitchell S, Waring P, Iezzi M, et al. Cutting edge: TRAIL deficiency accelerates hematological malignancies. J Immunol. 2005;175:5586–90.
Yue HH, Diehl GE, Winoto A. Loss of TRAIL-R does not affect thymic or intestinal tumor development in p53 and adenomatous polyposis coli mutant mice. Cell Death Differ. 2005;12:94–97.
LA OR, Tai L, Lee L, Kruse EA, Grabow S, Fairlie WD, et al. Membrane-bound Fas ligand only is essential for Fas-induced apoptosis. Nature. 2009;461:659–63.
Guillen-Ahlers H, Suckow MA, Castellino FJ, Ploplis VA. Fas/CD95 deficiency in ApcMin/+ mice increases intestinal tumor burden. PLoS ONE. 2010;5:e9070.
Park SM, Chen L, Zhang M, Ashton-Rickardt P, Turner JR, Peter ME. CD95 is cytoprotective for intestinal epithelial cells in colitis. Inflamm Bowel Dis. 2010;16:1063–70.
Fingleton B, Carter KJ, Matrisian LM. Loss of functional Fas ligand enhances intestinal tumorigenesis in the Min mouse model. Cancer Res. 2007;67:4800–6.
Kim JY, Kim YM, Park JM, Han YM, Lee KC, Hahm KB, et al. Cancer preventive effect of recombinant TRAIL by ablation of oncogenic inflammation in colitis-associated cancer rather than anticancer effect. Oncotarget. 2018;9:1705–16.
Lopetuso LR, Petito V, Zinicola T, Graziani C, Gerardi V, Arena V, et al. Infliximab does not increase colonic cancer risk associated to murine chronic colitis. World J Gastroenterol. 2016;22:9727–33.
Craven B, Zaric V, Martin A, Mureau C, Egan LJ. Effect of genetic deletion or pharmacological antagonism of tumor necrosis factor alpha on colitis-associated carcinogenesis in mice. Inflamm Bowel Dis. 2015;21:485–95.
Nyboe Andersen N, Pasternak B, Basit S, Andersson M, Svanström H, Caspersen S, et al. Association between tumor necrosis factor-α antagonists and risk of cancer in patients with inflammatory bowel disease. JAMA. 2014;311:2406–13.
Chang F, Lacey MR, Bouljihad M, Höner Zu Bentrup K, Fortgang IS. Tumor necrosis factor receptor 1 functions as a tumor suppressor. Am J Physiol Gastrointest Liver Physiol. 2012;302:G195–206.
Ba H, Jiang R, Zhang M, Yin B, Wang J, Li Z, et al. Suppression of transmembrane tumor necrosis factor alpha processing by a specific antibody protects against colitis-associated cancer. Front Immunol. 2021;12:687874.
Yang Y, Gharaibeh RZ, Newsome RC, Jobin C. Amending microbiota by targeting intestinal inflammation with TNF blockade attenuates development of colorectal cancer. Nat Cancer. 2020;1:723–34.
Kim YJ, Hong KS, Chung JW, Kim JH, Hahm KB. Prevention of colitis-associated carcinogenesis with infliximab. Cancer Prev Res. 2010;3:1314–33.
Onizawa M, Nagaishi T, Kanai T, Nagano K, Oshima S, Nemoto Y, et al. Signaling pathway via TNF-alpha/NF-kappaB in intestinal epithelial cells may be directly involved in colitis-associated carcinogenesis. Am J Physiol Gastrointest Liver Physiol. 2009;296:G850–859.
Rao VP, Poutahidis T, Ge Z, Nambiar PR, Horwitz BH, Fox JG, et al. Proinflammatory CD4+ CD45RB(hi) lymphocytes promote mammary and intestinal carcinogenesis in Apc(Min/+) mice. Cancer Res. 2006;66:57–61.
Oshima H, Ishikawa T, Yoshida GJ, Naoi K, Maeda Y, Naka K, et al. TNF-α/TNFR1 signaling promotes gastric tumorigenesis through induction of Noxo1 and Gna14 in tumor cells. Oncogene. 2014;33:3820–9.
Popivanova BK, Kitamura K, Wu Y, Kondo T, Kagaya T, Kaneko S, et al. Blocking TNF-alpha in mice reduces colorectal carcinogenesis associated with chronic colitis. J Clin Investig. 2008;118:560–70.
Castets M, Broutier L, Molin Y, Brevet M, Chazot G, Gadot N, et al. DCC constrains tumour progression via its dependence receptor activity. Nature. 2011;482:534–7.
Genevois AL, Ichim G, Coissieux MM, Lambert MP, Lavial F, Goldschneider D, et al. Dependence receptor TrkC is a putative colon cancer tumor suppressor. Proc Natl Acad Sci USA. 2013;110:3017–22.
Negulescu AM, Mehlen P. Dependence receptors - the dark side awakens. FEBS J. 2018;285:3909–24.
Grandin M, Meier M, Delcros JG, Nikodemus D, Reuten R, Patel TR, et al. Structural decoding of the Netrin-1/UNC5 interaction and its therapeutical implications in cancers. Cancer Cell. 2016;29:173–85.
Chen L, Park SM, Tumanov AV, Hau A, Sawada K, Feig C, et al. CD95 promotes tumour growth. Nature. 2010;465:492–6.
Pikarsky E, Porat RM, Stein I, Abramovitch R, Amit S, Kasem S, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004;431:461–6.
Schneider AT, Gautheron J, Feoktistova M, Roderburg C, Loosen SH, Roy S, et al. RIPK1 suppresses a TRAF2-dependent pathway to liver cancer. Cancer Cell. 2017;31:94–109.
Vucur M, Reisinger F, Gautheron J, Janssen J, Roderburg C, Cardenas DV, et al. RIP3 inhibits inflammatory hepatocarcinogenesis but promotes cholestasis by controlling caspase-8- and JNK-dependent compensatory cell proliferation. Cell Rep. 2013;4:776–90.
Vredevoogd DW, Kuilman T, Ligtenberg MA, Boshuizen J, Stecker KE, de Bruijn B, et al. Augmenting immunotherapy impact by lowering tumor TNF cytotoxicity threshold. Cell. 2019;178:585–599.e515.
Boege Y, Malehmir M, Healy ME, Bettermann K, Lorentzen A, Vucur M, et al. A dual role of caspase-8 in triggering and sensing proliferation-associated DNA damage, a key determinant of liver cancer development. Cancer Cell. 2017;32:342–359.e310.
Liccardi G, Ramos Garcia L, Tenev T, Annibaldi A, Legrand AJ, Robertson D, et al. RIPK1 and caspase-8 ensure chromosome stability independently of their role in cell death and inflammation. Mol Cell. 2019;73:413–428.e417.
Hakem A, El Ghamrasni S, Maire G, Lemmers B, Karaskova J, Jurisicova A, et al. Caspase-8 is essential for maintaining chromosomal stability and suppressing B-cell lymphomagenesis. Blood. 2012;119:3495–502.
Krelin Y, Zhang L, Kang TB, Appel E, Kovalenko A, Wallach D. Caspase-8 deficiency facilitates cellular transformation in vitro. Cell Death Differ. 2008;15:1350–5.
Rodriguez YI, Campos LE, Castro MG, Bannoud N, Blidner AG, Filippa VP, et al. Tumor necrosis factor receptor-1 (p55) deficiency attenuates tumor growth and intratumoral angiogenesis and stimulates CD8(+) T cell function in melanoma. Cells. 2020;9:249.
Schioppa T, Moore R, Thompson RG, Rosser EC, Kulbe H, Nedospasov S, et al. B regulatory cells and the tumor-promoting actions of TNF-α during squamous carcinogenesis. Proc Natl Acad Sci USA. 2011;108:10662–7.
Arnott CH, Scott KA, Moore RJ, Robinson SC, Thompson RG, Balkwill FR. Expression of both TNF-alpha receptor subtypes is essential for optimal skin tumour development. Oncogene. 2004;23:1902–10.
Scott KA, Moore RJ, Arnott CH, East N, Thompson RG, Scallon BJ, et al. An anti-tumor necrosis factor-alpha antibody inhibits the development of experimental skin tumors. Mol Cancer Ther. 2003;2:445–51.
Suganuma M, Okabe S, Marino MW, Sakai A, Sueoka E, Fujiki H. Essential role of tumor necrosis factor alpha (TNF-alpha) in tumor promotion as revealed by TNF-alpha-deficient mice. Cancer Res. 1999;59:4516–8.
Moore RJ, Owens DM, Stamp G, Arnott C, Burke F, East N, et al. Mice deficient in tumor necrosis factor-alpha are resistant to skin carcinogenesis. Nat Med. 1999;5:828–31.
Calıskan E, Gamsızkan M, Yurekli A, Botsali A, Kabalar ME, Demiriz M, et al. Anti-TNF agent etanercept augments UV-induced skin cancer development in SKH-1 mice. J Dermatol Treat. 2021;32:812–8.
Singh A, Singh A, Bauer SJ, Wheeler DL, Havighurst TC, Kim K, et al. Genetic deletion of TNFα inhibits ultraviolet radiation-induced development of cutaneous squamous cell carcinomas in PKCε transgenic mice via inhibition of cell survival signals. Carcinogenesis. 2016;37:72–80.
Lind MH, Rozell B, Wallin RP, van Hogerlinden M, Ljunggren HG, Toftgård R, et al. Tumor necrosis factor receptor 1-mediated signaling is required for skin cancer development induced by NF-kappaB inhibition. Proc Natl Acad Sci USA. 2004;101:4972–7.
Galheigo MR, Cruz AR, Cabral ÁS, Faria PR, Cordeiro RS, Silva MJ, et al. Role of the TNF-α receptor type 1 on prostate carcinogenesis in knockout mice. Prostate. 2016;76:917–26.
Sobo-Vujanovic A, Vujanovic L, DeLeo AB, Concha-Benavente F, Ferris RL, Lin Y, et al. Inhibition of Soluble tumor necrosis factor prevents chemically induced carcinogenesis in mice. Cancer Immunol Res. 2016;4:441–51.
He L, Bhat K, Duhacheck-Muggy S, Ioannidis A, Zhang L, Nguyen NT, et al. Tumor necrosis factor receptor signaling modulates carcinogenesis in a mouse model of breast cancer. Neoplasia. 2021;23:197–209.
Sangaletti S, Tripodo C, Ratti C, Piconese S, Porcasi R, Salcedo R, et al. Oncogene-driven intrinsic inflammation induces leukocyte production of tumor necrosis factor that critically contributes to mammary carcinogenesis. Cancer Res. 2010;70:7764–75.
Chadwick JW, Macdonald R, Ali AA, Glogauer M, Magalhaes MA. TNFα signaling is increased in progressing oral potentially malignant disorders and regulates malignant transformation in an oral carcinogenesis model. Front Oncol. 2021;11:741013.
Karabela SP, Kairi CA, Magkouta S, Psallidas I, Moschos C, Stathopoulos I, et al. Neutralization of tumor necrosis factor bioactivity ameliorates urethane-induced pulmonary oncogenesis in mice. Neoplasia. 2011;13:1143–51.
Gong L, da Silva Caetano M, Cumpian AM, Daliri S, Garza Flores A, Chang SH, et al. Tumor necrosis factor links chronic obstructive pulmonary disease and K-ras mutant lung cancer through induction of an immunosuppressive pro-tumor microenvironment. Oncoimmunology. 2016;5:e1229724.
Kedinger V, Muller S, Gronemeyer H. Targeted expression of tumor necrosis factor-related apoptosis-inducing ligand TRAIL in skin protects mice against chemical carcinogenesis. Mol Cancer. 2011;10:34.
Chio II, Sasaki M, Ghazarian D, Moreno J, Done S, Ueda T, et al. TRADD contributes to tumour suppression by regulating ULF-dependent p19Arf ubiquitylation. Nat Cell Biol. 2012;14:625–33.
Grosse-Wilde A, Voloshanenko O, Bailey SL, Longton GM, Schaefer U, Csernok AI, et al. TRAIL-R deficiency in mice enhances lymph node metastasis without affecting primary tumor development. J Clin Investig. 2008;118:100–10.
Takeda K, Smyth MJ, Cretney E, Hayakawa Y, Kayagaki N, Yagita H, et al. Critical role for tumor necrosis factor-related apoptosis-inducing ligand in immune surveillance against tumor development. J Exp Med. 2002;195:161–9.
Cretney E, Takeda K, Yagita H, Glaccum M, Peschon JJ, Smyth MJ. Increased susceptibility to tumor initiation and metastasis in TNF-related apoptosis-inducing ligand-deficient mice. J Immunol. 2002;168:1356–61.
Montinaro A, Areso Zubiaur I, Saggau J, Kretz AL, Ferreira RMM, Hassan O, et al. Potent pro-apoptotic combination therapy is highly effective in a broad range of cancers. Cell Death Differ. 2022;29:492–503.
von Karstedt S, Conti A, Nobis M, Montinaro A, Hartwig T, Lemke J, et al. Cancer cell-autonomous TRAIL-R signaling promotes KRAS-driven cancer progression, invasion, and metastasis. Cancer Cell. 2015;27:561–73.
Hartwig T, Montinaro A, von Karstedt S, Sevko A, Surinova S, Chakravarthy A, et al. The TRAIL-induced cancer secretome promotes a tumor-supportive immune microenvironment via CCR2. Mol Cell. 2017;65:730–742.e735.
Hoogwater FJ, Nijkamp MW, Smakman N, Steller EJ, Emmink BL, Westendorp BF, et al. Oncogenic K-Ras turns death receptors into metastasis-promoting receptors in human and mouse colorectal cancer cells. Gastroenterology. 2010;138:2357–67.
Rieux-Laucat F, Le Deist F, Hivroz C, Roberts IA, Debatin KM, Fischer A, et al. Mutations in Fas associated with human lymphoproliferative syndrome and autoimmunity. Science. 1995;268:1347–9.
Watanabe-Fukunaga R, Brannan CI, Copeland NG, Jenkins NA, Nagata S. Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediates apoptosis. Nature. 1992;356:314–7.
Alvarez-Diaz S, Dillon CP, Lalaoui N, Tanzer MC, Rodriguez DA, Lin A, et al. The pseudokinase MLKL and the kinase RIPK3 have distinct roles in autoimmune disease caused by loss of death-receptor-induced apoptosis. Immunity. 2016;45:513–26.
Chyuan IT, Tsai HF, Wu CS, Sung CC, Hsu PN. TRAIL-mediated suppression of T cell receptor signaling inhibits T cell activation and inflammation in experimental autoimmune encephalomyelitis. Front Immunol. 2018;9:15.
Ikeda T, Hirata S, Fukushima S, Matsunaga Y, Ito T, Uchino M, et al. Dual effects of TRAIL in suppression of autoimmunity: the inhibition of Th1 cells and the promotion of regulatory T cells. J Immunol. 2010;185:5259–67.
Cretney E, McQualter JL, Kayagaki N, Yagita H, Bernard CC, Grewal IS, et al. TNF-related apoptosis-inducing ligand (TRAIL)/Apo2L suppresses experimental autoimmune encephalomyelitis in mice. Immunol cell Biol. 2005;83:511–9.
Razmara M, Hilliard B, Ziarani AK, Murali R, Yellayi S, Ghazanfar M, et al. Fn14-TRAIL, a chimeric intercellular signal exchanger, attenuates experimental autoimmune encephalomyelitis. Am J Pathol. 2009;174:460–74.
Aktas O, Smorodchenko A, Brocke S, Infante-Duarte C, Schulze Topphoff U, Vogt J, et al. Neuronal damage in autoimmune neuroinflammation mediated by the death ligand TRAIL. Neuron. 2005;46:421–32.
Hilliard B, Wilmen A, Seidel C, Liu TS, Göke R, Chen Y. Roles of TNF-related apoptosis-inducing ligand in experimental autoimmune encephalomyelitis. J Immunol. 2001;166:1314–9.
Lamhamedi-Cherradi SE, Zheng SJ, Maguschak KA, Peschon J, Chen YH. Defective thymocyte apoptosis and accelerated autoimmune diseases in TRAIL−/− mice. Nat Immunol. 2003;4:255–60.
Song K, Chen Y, Göke R, Wilmen A, Seidel C, Göke A, et al. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is an inhibitor of autoimmune inflammation and cell cycle progression. J Exp Med. 2000;191:1095–104.
Park JS, Oh Y, Park O, Foss CA, Lim SM, Jo DG, et al. PEGylated TRAIL ameliorates experimental inflammatory arthritis by regulation of Th17 cells and regulatory T cells. J Control Release. 2017;267:163–71.
Chyuan IT, Tsai HF, Liao HJ, Wu CS, Hsu PN. An apoptosis-independent role of TRAIL in suppressing joint inflammation and inhibiting T-cell activation in inflammatory arthritis. Cell Mol Immunol. 2018;15:846–57.
Jin CH, Chae SY, Kim TH, Yang HK, Lee EY, Song YW, et al. Effect of tumor necrosis factor-related apoptosis-inducing ligand on the reduction of joint inflammation in experimental rheumatoid arthritis. J Pharmacol Exp Ther. 2010;332:858–65.
Kang S, Park EJ, Joe Y, Seo E, Park MK, Seo SY, et al. Systemic delivery of TNF-related apoptosis-inducing ligand (TRAIL) elevates levels of tissue inhibitor of metalloproteinase-1 (TIMP-1) and prevents type 1 diabetes in nonobese diabetic mice. Endocrinology. 2010;151:5638–46.
Mi QS, Ly D, Lamhamedi-Cherradi SE, Salojin KV, Zhou L, Grattan M, et al. Blockade of tumor necrosis factor-related apoptosis-inducing ligand exacerbates type 1 diabetes in NOD mice. Diabetes. 2003;52:1967–75.
Bossi F, Bernardi S, Zauli G, Secchiero P, Fabris B. TRAIL modulates the immune system and protects against the development of diabetes. J Immunol Res. 2015;2015:680749.
Lamhamedi-Cherradi SE, Zheng S, Tisch RM, Chen YH. Critical roles of tumor necrosis factor-related apoptosis-inducing ligand in type 1 diabetes. Diabetes. 2003;52:2274–8.
Bachmann R, Eugster HP, Frei K, Fontana A, Lassmann H. Impairment of TNF-receptor-1 signaling but not fas signaling diminishes T-cell apoptosis in myelin oligodendrocyte glycoprotein peptide-induced chronic demyelinating autoimmune encephalomyelitis in mice. Am J Pathol. 1999;154:1417–22.
Malipiero U, Frei K, Spanaus KS, Agresti C, Lassmann H, Hahne M, et al. Myelin oligodendrocyte glycoprotein-induced autoimmune encephalomyelitis is chronic/relapsing in perforin knockout mice, but monophasic in Fas- and Fas ligand-deficient lpr and gld mice. Eur J Immunol. 1997;27:3151–60.
Waldner H, Sobel RA, Howard E, Kuchroo VK. Fas- and FasL-deficient mice are resistant to induction of autoimmune encephalomyelitis. J Immunol. 1997;159:3100–3.
Sabelko KA, Kelly KA, Nahm MH, Cross AH, Russell JH. Fas and Fas ligand enhance the pathogenesis of experimental allergic encephalomyelitis, but are not essential for immune privilege in the central nervous system. J Immunol. 1997;159:3096–9.
Wolf Y, Shemer A, Polonsky M, Gross M, Mildner A, Yona S, et al. Autonomous TNF is critical for in vivo monocyte survival in steady state and inflammation. J Exp Med. 2017;214:905–17.
Williams SK, Fairless R, Maier O, Liermann PC, Pichi K, Fischer R, et al. Anti-TNFR1 targeting in humanized mice ameliorates disease in a model of multiple sclerosis. Sci Rep. 2018;8:13628.
Williams SK, Maier O, Fischer R, Fairless R, Hochmeister S, Stojic A, et al. Antibody-mediated inhibition of TNFR1 attenuates disease in a mouse model of multiple sclerosis. PLoS ONE. 2014;9:e90117.
Nomura T, Abe Y, Kamada H, Shibata H, Kayamuro H, Inoue M, et al. Therapeutic effect of PEGylated TNFR1-selective antagonistic mutant TNF in experimental autoimmune encephalomyelitis mice. J Control Release. 2011;149:8–14.
Steeland S, Van Ryckeghem S, Van Imschoot G, De Rycke R, Toussaint W, Vanhoutte L, et al. TNFR1 inhibition with a Nanobody protects against EAE development in mice. Sci Rep. 2017;7:13646.
Brambilla R, Ashbaugh JJ, Magliozzi R, Dellarole A, Karmally S, Szymkowski DE, et al. Inhibition of soluble tumour necrosis factor is therapeutic in experimental autoimmune encephalomyelitis and promotes axon preservation and remyelination. Brain. 2011;134:2736–54.
Körner H, Lemckert FA, Chaudhri G, Etteldorf S, Sedgwick JD. Tumor necrosis factor blockade in actively induced experimental autoimmune encephalomyelitis prevents clinical disease despite activated T cell infiltration to the central nervous system. Eur J Immunol. 1997;27:1973–81.
Körner H, Goodsall AL, Lemckert FA, Scallon BJ, Ghrayeb J, Ford AL, et al. Unimpaired autoreactive T-cell traffic within the central nervous system during tumor necrosis factor receptor-mediated inhibition of experimental autoimmune encephalomyelitis. Proc Natl Acad Sci USA. 1995;92:11066–70.
Richter F, Williams SK, John K, Huber C, Vaslin C, Zanker H, et al. The TNFR1 antagonist atrosimab is therapeutic in mouse models of acute and chronic inflammation. Front Immunol. 2021;12:705485.
Dittel BN, Merchant RM, Janeway CA Jr. Evidence for Fas-dependent and Fas-independent mechanisms in the pathogenesis of experimental autoimmune encephalomyelitis. J Immunol. 1999;162:6392–6400.
Suvannavejh GC, Dal Canto MC, Matis LA, Miller SD. Fas-mediated apoptosis in clinical remissions of relapsing experimental autoimmune encephalomyelitis. J Clin Investig. 2000;105:223–31.
Wang X, Haroon F, Karray S, Martina D, Schlüter D. Astrocytic Fas ligand expression is required to induce T-cell apoptosis and recovery from experimental autoimmune encephalomyelitis. Eur J Immunol. 2013;43:115–24.
Sabelko-Downes KA, Cross AH, Russell JH. Dual role for Fas ligand in the initiation of and recovery from experimental allergic encephalomyelitis. J Exp Med. 1999;189:1195–205.
Batoulis H, Recks MS, Holland FO, Thomalla F, Williams RO, Kuerten S. Blockade of tumour necrosis factor-α in experimental autoimmune encephalomyelitis reveals differential effects on the antigen-specific immune response and central nervous system histopathology. Clin Exp Immunol. 2014;175:41–48.
Liu J, Marino MW, Wong G, Grail D, Dunn A, Bettadapura J, et al. TNF is a potent anti-inflammatory cytokine in autoimmune-mediated demyelination. Nat Med. 1998;4:78–83.
Tu-Rapp H, Hammermüller A, Mix E, Kreutzer HJ, Goerlich R, Köhler H, et al. A proinflammatory role for Fas in joints of mice with collagen-induced arthritis. Arthritis Res Ther. 2004;6:R404–14.
Shen F, Verma AH, Volk A, Jones B, Coleman BM, Loza MJ, et al. Combined blockade of TNF-α and IL-17A alleviates progression of collagen-induced arthritis without causing serious infections in mice. J Immunol. 2019;202:2017–26.
Moore AR, Allden S, Bourne T, Denis MC, Kranidioti K, Okoye R, et al. Collagen II antibody-induced arthritis in Tg1278TNFko mice: optimization of a novel model to assess treatments targeting human TNFα in rheumatoid arthritis. J Transl Med. 2014;12:285.
Zalevsky J, Secher T, Ezhevsky SA, Janot L, Steed PM, O’Brien C, et al. Dominant-negative inhibitors of soluble TNF attenuate experimental arthritis without suppressing innate immunity to infection. J Immunol. 2007;179:1872–83.
Zhao Y, Yang X, Li S, Zhang B, Li S, Wang X, et al. sTNFRII-Fc modification protects human UC-MSCs against apoptosis/autophagy induced by TNF-α and enhances their efficacy in alleviating inflammatory arthritis. Stem Cell Res Ther. 2021;12:535.
Huang QQ, Birkett R, Koessler RE, Cuda CM, Haines GK 3rd, Jin JP, et al. Fas signaling in macrophages promotes chronicity in K/BxN serum-induced arthritis. Arthritis Rheumatol. 2014;66:68–77.
Kang SE, Park JK, Yoo HJ, Kang HS, Park YW, Park BC, et al. Efficacy of novel bispecific antibody targeting TNF-α/CXCL10 in the treatment of experimental arthritis. Transl Res. 2021;232:75–87.
Itoh N, Imagawa A, Hanafusa T, Waguri M, Yamamoto K, Iwahashi H, et al. Requirement of Fas for the development of autoimmune diabetes in nonobese diabetic mice. J Exp Med. 1997;186:613–8.
Su X, Hu Q, Kristan JM, Costa C, Shen Y, Gero D, et al. Significant role for Fas in the pathogenesis of autoimmune diabetes. J Immunol. 2000;164:2523–32.
Chervonsky AV, Wang Y, Wong FS, Visintin I, Flavell RA, Janeway CA Jr, et al. The role of Fas in autoimmune diabetes. Cell. 1997;89:17–24.
Vence L, Benoist C, Mathis D. Fas deficiency prevents type 1 diabetes by inducing hyporesponsiveness in islet beta-cell-reactive T-cells. Diabetes. 2004;53:2797–803.
Mohamood AS, Guler ML, Xiao Z, Zheng D, Hess A, Wang Y, et al. Protection from autoimmune diabetes and T-cell lymphoproliferation induced by FasL mutation are differentially regulated and can be uncoupled pharmacologically. Am J Pathol. 2007;171:97–106.
Jeong JH, Kim SH, Lee M, Kim WJ, Park TG, Ko KS, et al. Non-viral systemic delivery of Fas siRNA suppresses cyclophosphamide-induced diabetes in NOD mice. J Control Release. 2010;143:88–94.
Trivedi PM, Fynch S, Kennedy LM, Chee J, Krishnamurthy B, O’Reilly LA, et al. Soluble FAS ligand is not required for pancreatic islet inflammation or beta-cell destruction in non-obese diabetic mice. Cell Death Discov. 2019;5:136.
Choi D, Radziszewska A, Schroer SA, Liadis N, Liu Y, Zhang Y, et al. Deletion of Fas in the pancreatic beta-cells leads to enhanced insulin secretion. Am J Physiol Endocrinol Metab. 2009;297:E1304–12.
Thomas HE, Darwiche R, Corbett JA, Kay TW. Evidence that beta cell death in the nonobese diabetic mouse is Fas independent. J Immunol. 1999;163:1562–9.
Biemans VBC, Sleutjes JAM, de Vries AC, Bodelier AGL, Dijkstra G, Oldenburg B, et al. Tofacitinib for ulcerative colitis: results of the prospective Dutch Initiative on Crohn and Colitis (ICC) registry. Aliment Pharmacol Ther. 2020;51:880–8.
Almon E, Shaaltiel Y, Sbeit W, Fich A, Schwartz D, Waterman M, et al. Novel orally administered recombinant anti-TNF alpha fusion protein for the treatment of ulcerative colitis: results from a phase 2a clinical trial. J Clin Gastroenterol. 2021;55:134–40.
Liu CY, Tam SS, Huang Y, Dubé PE, Alhosh R, Girish N, et al. TNF receptor 1 promotes early-life immunity and protects against colitis in mice. Cell Rep. 2020;33:108275.
Lin DP, Jin YL, Hu DY, Ying SJ, Jiang Y. Influence of TRAIL deficiency on Th17 cells and colonic microbiota in experimental colitis mouse model. Am J Med Sci. 2021;362:188–97.
Chyuan IT, Tsai HF, Wu CS, Hsu PN. TRAIL suppresses gut inflammation and inhibits colitogeic T-cell activation in experimental colitis via an apoptosis-independent pathway. Mucosal Immunol. 2019;12:980–9.
Pinhu L, Qin Y, Xiong B, You Y, Li J, Sooranna SR. Overexpression of Fas and FasL is associated with infectious complications and severity of experimental severe acute pancreatitis by promoting apoptosis of lymphocytes. Inflammation. 2014;37:1202–12.
Randhi R, Damon M, Dixon KJ. Selective inhibition of soluble TNF using XPro1595 relieves pain and attenuates cerulein-induced pathology in mice. BMC Gastroenterol. 2021;21:243.
Mao XJ, Zhang XM, Zhang HL, Quezada HC, Mix E, Yang X, et al. TNF-alpha receptor 1 deficiency reduces antigen-presenting capacity of Schwann cells and ameliorates experimental autoimmune neuritis in mice. Neurosci Lett. 2010;470:19–23.
Taylor JM, Pollard JD. Soluble TNFR1 inhibits the development of experimental autoimmune neuritis by modulating blood-nerve-barrier permeability and inflammation. J Neuroimmunol. 2007;183:118–24.
Bao L, Lindgren JU, Zhu Y, Ljunggren HG, Zhu J. Exogenous soluble tumor necrosis factor receptor type I ameliorates murine experimental autoimmune neuritis. Neurobiol Dis. 2003;12:73–81.
Lu MO, Duan RS, Quezada HC, Chen ZG, Mix E, Jin T, et al. Aggravation of experimental autoimmune neuritis in TNF-alpha receptor 1 deficient mice. J Neuroimmunol. 2007;186:19–26.
Kaaij MH, Rip J, Jeucken KCM, Kan YY, van Rooijen CCN, Saris J, et al. Overexpression of transmembrane TNF drives development of ectopic lymphoid structures in the bone marrow and B cell lineage alterations in experimental spondyloarthritis. J Immunol. 2021;207:2337–46.
Chen S, Lin Z, Xi L, Zheng Y, Zhou Q, Chen X. Differential role of TNFR1 and TNFR2 in the development of imiquimod-induced mouse psoriasis. J Leukoc Biol. 2021;110:1047–55.
Yu X, Li L, Li Q, Zang X, Liu Z. TRAIL and DR5 promote thyroid follicular cell apoptosis in iodine excess-induced experimental autoimmune thyroiditis in NOD mice. Biol Trace Elem Res. 2011;143:1064–76.
Fang Y, Sharp GC, Yagita H, Braley-Mullen H. A critical role for TRAIL in resolution of granulomatous experimental autoimmune thyroiditis. J Pathol. 2008;216:505–13.
Wei Y, Chen K, Sharp GC, Braley-Mullen H. Fas ligand is required for resolution of granulomatous experimental autoimmune thyroiditis. J Immunol. 2004;173:7615–21.
Wang SH, Chen GH, Fan Y, Van Antwerp M, Baker JR Jr. Tumor necrosis factor-related apoptosis-inducing ligand inhibits experimental autoimmune thyroiditis by the expansion of CD4+CD25+ regulatory T cells. Endocrinology. 2009;150:2000–7.
Wang SH, Cao Z, Wolf JM, Van Antwerp M, Baker JR Jr. Death ligand tumor necrosis factor-related apoptosis-inducing ligand inhibits experimental autoimmune thyroiditis. Endocrinology. 2005;146:4721–6.
Patankar JV, Müller TM, Kantham S, Acera MG, Mascia F, Scheibe K, et al. E-type prostanoid receptor 4 drives resolution of intestinal inflammation by blocking epithelial necroptosis. Nat Cell Biol. 2021;23:796–807.
Kang TB, Jeong JS, Yang SH, Kovalenko A, Wallach D. Caspase-8 deficiency in mouse embryos triggers chronic RIPK1-dependent activation of inflammatory genes, independently of RIPK3. Cell Death Differ. 2018;25:1107–17.
Rajput A, Kovalenko A, Bogdanov K, Yang SH, Kang TB, Kim JC, et al. RIG-I RNA helicase activation of IRF3 transcription factor is negatively regulated by caspase-8-mediated cleavage of the RIP1 protein. Immunity. 2011;34:340–51.
Laurien L, Nagata M, Schünke H, Delanghe T, Wiederstein JL, Kumari S, et al. Autophosphorylation at serine 166 regulates RIP kinase 1-mediated cell death and inflammation. Nat Commun. 2020;11:1747.
Rickard JA, Anderton H, Etemadi N, Nachbur U, Darding M, Peltzer N, et al. TNFR1-dependent cell death drives inflammation in Sharpin-deficient mice. eLife 2014;3:e03464.
Kumari S, Redouane Y, Lopez-Mosqueda J, Shiraishi R, Romanowska M, Lutzmayer S, et al. Sharpin prevents skin inflammation by inhibiting TNFR1-induced keratinocyte apoptosis. eLife. 2014;3:e03422.
Berger SB, Kasparcova V, Hoffman S, Swift B, Dare L, Schaeffer M, et al. Cutting Edge: RIP1 kinase activity is dispensable for normal development but is a key regulator of inflammation in SHARPIN-deficient mice. J Immunol. 2014;192:5476–80.
Taraborrelli L, Peltzer N, Montinaro A, Kupka S, Rieser E, Hartwig T, et al. LUBAC prevents lethal dermatitis by inhibiting cell death induced by TNF, TRAIL and CD95L. Nat Commun. 2018;9:3910.
Mc Guire C, Volckaert T, Wolke U, Sze M, de Rycke R, Waisman A, et al. Oligodendrocyte-specific FADD deletion protects mice from autoimmune-mediated demyelination. J Immunol. 2010;185:7646–53.
Sun J, Hilliard B, Xu L, Chen YH. Essential roles of the Fas-associated death domain in autoimmune encephalomyelitis. J Immunol. 2005;175:4783–8.
Newton K, Harris AW, Bath ML, Smith KG, Strasser A. A dominant interfering mutant of FADD/MORT1 enhances deletion of autoreactive thymocytes and inhibits proliferation of mature T lymphocytes. EMBO J. 1998;17:706–18.
Zhang CJ, Jiang M, Zhou H, Liu W, Wang C, Kang Z, et al. TLR-stimulated IRAKM activates caspase-8 inflammasome in microglia and promotes neuroinflammation. J Clin Investig. 2018;128:5399–412.
Allison J, Thomas HE, Catterall T, Kay TW, Strasser A. Transgenic expression of dominant-negative Fas-associated death domain protein in beta cells protects against Fas ligand-induced apoptosis and reduces spontaneous diabetes in nonobese diabetic mice. J Immunol. 2005;175:293–301.
Mollah ZU, Wali J, McKenzie MD, Krishnamurthy B, Graham KL, Fynch S, et al. The pro-apoptotic BH3-only protein Bid is dispensable for development of insulitis and diabetes in the non-obese diabetic mouse. Apoptosis. 2011;16:822–30.
Huang QQ, Birkett R, Doyle RE, Haines GK, Perlman H, Shi B, et al. Association of increased F4/80(high) macrophages with suppression of serum-transfer arthritis in mice with reduced FLIP in myeloid cells. Arthritis Rheumatol. 2017;69:1762–71.
Dominguez S, Montgomery AB, Haines GK 3rd, Bloomfield CL, Cuda CM. The caspase-8/RIPK3 signaling axis in antigen presenting cells controls the inflammatory arthritic response. Arthritis Res Ther. 2017;19:224.
Pearson JS, Giogha C, Ong SY, Kennedy CL, Kelly M, Robinson KS, et al. A type III effector antagonizes death receptor signalling during bacterial gut infection. Nature. 2013;501:247–51.
Li S, Zhang L, Yao Q, Li L, Dong N, Rong J, et al. Pathogen blocks host death receptor signalling by arginine GlcNAcylation of death domains. Nature 2013;501:242–6.
Uchiyama R, Yonehara S, Taniguchi S, Ishido S, Ishii KJ, Tsutsui H. Inflammasome and Fas-mediated IL-1β contributes to Th17/Th1 cell induction in pathogenic bacterial infection in vivo. J Immunol. 2017;199:1122–30.
Maudet C, Kheloufi M, Levallois S, Gaillard J, Huang L, Gaultier C, et al. Bacterial inhibition of Fas-mediated killing promotes neuroinvasion and persistence. Nature. 2022;603:900–6.
Krzyzowska M, Baska P, Orlowski P, Zdanowski R, Winnicka A, Eriksson K, et al. HSV-2 regulates monocyte inflammatory response via the Fas/FasL pathway. PLoS ONE. 2013;8:e70308.
O’Donnell JA, Kennedy CL, Pellegrini M, Nowell CJ, Zhang JG, O’Reilly LA, et al. Fas regulates neutrophil lifespan during viral and bacterial infection. J Leukoc Biol. 2015;97:321–6.
Peterson LW, Philip NH, DeLaney A, Wynosky-Dolfi MA, Asklof K, Gray F, et al. RIPK1-dependent apoptosis bypasses pathogen blockade of innate signaling to promote immune defense. J Exp Med. 2017;214:3171–82.
Weng D, Marty-Roix R, Ganesan S, Proulx MK, Vladimer GI, Kaiser WJ, et al. Caspase-8 and RIP kinases regulate bacteria-induced innate immune responses and cell death. Proc Natl Acad Sci USA. 2014;111:7391–6.
DeLaney AA, Berry CT, Christian DA, Hart A, Bjanes E, Wynosky-Dolfi MA, et al. Caspase-8 promotes c-Rel-dependent inflammatory cytokine expression and resistance against Toxoplasma gondii. Proc Natl Acad Sci USA. 2019;116:11926–35.
Kuriakose T, Man SM, Malireddi RK, Karki R, Kesavardhana S, Place DE, et al. ZBP1/DAI is an innate sensor of influenza virus triggering the NLRP3 inflammasome and programmed cell death pathways. Sci Immunol. 2016;1:aag2045.
Thapa RJ, Ingram JP, Ragan KB, Nogusa S, Boyd DF, Benitez AA, et al. DAI senses influenza A virus genomic RNA and activates RIPK3-dependent cell death. Cell Host Microbe. 2016;20:674–81.
Oltean T, Van San E, Divert T, Vanden Berghe T, Saelens X, Maelfait J, et al. Viral dosing of influenza A infection reveals involvement of RIPK3 and FADD, but not MLKL. Cell Death Dis. 2021;12:471.
Nogusa S, Thapa RJ, Dillon CP, Liedmann S, Oguin TH, 3rd, Ingram JP, et al. RIPK3 activates parallel pathways of mlkl-driven necroptosis and fadd-mediated apoptosis to protect against influenza a virus. Cell Host Microbe 2016;20:13–24.
Zhang T, Yin C, Boyd DF, Quarato G, Ingram JP, Shubina M, et al. influenza virus z-rnas induce zbp1-mediated necroptosis. Cell 2020;180:1115–29.e1113.
Place DE, Christgen S, Tuladhar S, Vogel P, Malireddi RKS, Kanneganti TD. Hierarchical cell death program disrupts the intracellular niche required for Burkholderia thailandensis pathogenesis. mBio. 2021;12:e0105921.
Ebert G, Preston S, Allison C, Cooney J, Toe JG, Stutz MD, et al. Cellular inhibitor of apoptosis proteins prevent clearance of hepatitis B virus. Proc Natl Acad Sci USA. 2015;112:5797–802.
Ebert G, Allison C, Preston S, Cooney J, Toe JG, Stutz MD, et al. Eliminating hepatitis B by antagonizing cellular inhibitors of apoptosis. Proc Natl Acad Sci USA. 2015;112:5803–8.
Ebert G, Lopaticki S, O'Neill MT, Steel RWJ, Doerflinger M, Rajasekaran P, et al. targeting the extrinsic pathway of hepatocyte apoptosis promotes clearance of plasmodium liver infection. Cell Rep. 2020;30:4343-4354.e4344.
Alikhani M, Alikhani Z, He H, Liu R, Popek BI, Graves DT. Lipopolysaccharides indirectly stimulate apoptosis and global induction of apoptotic genes in fibroblasts. J Biol Chem. 2003;278:52901–8.
Sarid R, Ben-Moshe T, Kazimirsky G, Weisberg S, Appel E, Kobiler D, et al. vFLIP protects PC-12 cells from apoptosis induced by Sindbis virus: implications for the role of TNF-alpha. Cell Death Differ. 2001;8:1224–31.
Qian Z, Shuying W, Ranran D. Inhibitory effects of JQ1 on listeria monocytogenes-induced acute liver injury by blocking BRD4/RIPK1 axis. Biomed Pharmacother. 2020;125:109818.
Kim H, Rhee SH, Pothoulakis C, Lamont JT. Inflammation and apoptosis in Clostridium difficile enteritis is mediated by PGE2 up-regulation of Fas ligand. Gastroenterology. 2007;133:875–86.
He BL, Yuan JM, Yang LY, Xie JF, Weng SP, Yu XQ, et al. The viral TRAF protein (ORF111L) from infectious spleen and kidney necrosis virus interacts with TRADD and induces caspase 8-mediated apoptosis. PLoS ONE. 2012;7:e37001.
Mandal P, Feng Y, Lyons JD, Berger SB, Otani S, DeLaney A, et al. Caspase-8 collaborates with caspase-11 to drive tissue damage and execution of endotoxic shock. Immunity. 2018;49:42–55.e46.
Karki R, Sharma BR, Tuladhar S, Williams EP, Zalduondo L, Samir P, et al. Synergism of TNF-α and IFN-γ triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell. 2021;184:149–168.e117.
Langen RC, Van Der Velden JL, Schols AM, Kelders MC, Wouters EF, Janssen-Heininger YM. Tumor necrosis factor-alpha inhibits myogenic differentiation through MyoD protein destabilization. FASEB J. 2004;18:227–37.
Kim D, Singh N, Waldemer-Streyer RJ, Yoon MS, Chen J. Muscle-derived TRAIL negatively regulates myogenic differentiation. Exp Cell Res. 2020;394:112165.
Zhang R, Wang L, He L, Yang B, Yao C, Du P, et al. Fas-associated protein with death domain regulates notch signaling during muscle regeneration. Cells Tissues Organs. 2014;200:253–64.
Chen SE, Jin B, Li YP. TNF-alpha regulates myogenesis and muscle regeneration by activating p38 MAPK. Am J Physiol Cell Physiol. 2007;292:C1660–1671.
Chen SE, Gerken E, Zhang Y, Zhan M, Mohan RK, Li AS, et al. Role of TNF-{alpha} signaling in regeneration of cardiotoxin-injured muscle. Am J Physiol Cell Physiol. 2005;289:C1179–1187.
Dufresne SS, Boulanger-Piette A, Bossé S, Argaw A, Hamoudi D, Marcadet L, et al. Genetic deletion of muscle RANK or selective inhibition of RANKL is not as effective as full-length OPG-fc in mitigating muscular dystrophy. Acta Neuropathol Commun. 2018;6:31.
Alger HM, Raben N, Pistilli E, Francia DL, Rawat R, Getnet D, et al. The role of TRAIL in mediating autophagy in myositis skeletal muscle: a potential nonimmune mechanism of muscle damage. Arthritis Rheum. 2011;63:3448–57.
Kondo M, Murakawa Y, Harashima N, Kobayashi S, Yamaguchi S, Harada M. Roles of proinflammatory cytokines and the Fas/Fas ligand interaction in the pathogenesis of inflammatory myopathies. Immunology. 2009;128:e589–99.
Del Sorbo L, Costamagna A, Muraca G, Rotondo G, Civiletti F, Vizio B, et al. Intratracheal Administration of Small Interfering RNA Targeting Fas Reduces Lung Ischemia-Reperfusion Injury. Crit Care Med. 2016;44:e604–13.
An S, Hishikawa Y, Liu J, Koji T. Lung injury after ischemia-reperfusion of small intestine in rats involves apoptosis of type II alveolar epithelial cells mediated by TNF-alpha and activation of Bid pathway. Apoptosis. 2007;12:1989–2001.
Patel BV, Wilson MR, O’Dea KP, Takata M. TNF-induced death signaling triggers alveolar epithelial dysfunction in acute lung injury. J Immunol. 2013;190:4274–82.
Wilson MR, Wakabayashi K, Bertok S, Oakley CM, Patel BV, O’Dea KP, et al. Inhibition of TNF Receptor p55 By a Domain Antibody Attenuates the Initial Phase of Acid-Induced Lung Injury in Mice. Front Immunol. 2017;8:128.
Bohr A, Tsapis N, Foged C, Andreana I, Yang M, Fattal E. Treatment of acute lung inflammation by pulmonary delivery of anti-TNF-α siRNA with PAMAM dendrimers in a murine model. Eur J Pharm Biopharm. 2020;156:114–20.
Lai WY, Wang JW, Huang BT, Lin EP, Yang PC. A novel TNF-α-targeting aptamer for TNF-α-mediated acute lung injury and acute liver failure. Theranostics. 2019;9:1741–51.
Proudfoot A, Bayliffe A, O’Kane CM, Wright T, Serone A, Bareille PJ, et al. Novel anti-tumour necrosis factor receptor-1 (TNFR1) domain antibody prevents pulmonary inflammation in experimental acute lung injury. Thorax. 2018;73:723–30.
Bohr A, Tsapis N, Andreana I, Chamarat A, Foged C, Delomenie C, et al. Anti-inflammatory effect of anti-TNF-α SiRNA cationic phosphorus dendrimer nanocomplexes administered intranasally in a murine acute lung injury model. Biomacromolecules. 2017;18:2379–88.
Weifeng Y, Li L, Yujie H, Weifeng L, Zhenhui G, Wenjie H. Inhibition of acute lung injury by TNFR-Fc through regulation of an inflammation-oxidative stress pathway. PLoS ONE. 2016;11:e0151672.
Cakarova L, Marsh LM, Wilhelm J, Mayer K, Grimminger F, Seeger W, et al. Macrophage tumor necrosis factor-alpha induces epithelial expression of granulocyte-macrophage colony-stimulating factor: impact on alveolar epithelial repair. Am J Respir Crit Care Med. 2009;180:521–32.
Matute-Bello G, Winn RK, Martin TR, Liles WC. Sustained lipopolysaccharide-induced lung inflammation in mice is attenuated by functional deficiency of the Fas/Fas ligand system. Clin Diagn Lab Immunol. 2004;11:358–61.
Janssen WJ, Barthel L, Muldrow A, Oberley-Deegan RE, Kearns MT, Jakubzick C, et al. Fas determines differential fates of resident and recruited macrophages during resolution of acute lung injury. Am J Respir Crit Care Med. 2011;184:547–60.
Qian L, Yin X, Ji J, Chen Z, Fang H, Li H, et al. Tumor necrosis factor-α small interfering RNA alveolar epithelial cell-targeting nanoparticles reduce lung injury in C57BL/6J mice with sepsis. J Int Med Res. 2021;49:300060520984652.
Weckbach S, Hohmann C, Denk S, Kellermann P, Huber-Lang MS, Baumann B, et al. Apoptotic and inflammatory signaling via Fas and tumor necrosis factor receptor I contribute to the development of chest trauma-induced septic acute lung injury. J Trauma Acute Care Surg. 2013;74:792–800.
Thakkar RK, Chung CS, Chen Y, Monaghan SF, Lomas-Neira J, Heffernan DS, et al. Local tissue expression of the cell death ligand, fas ligand, plays a central role in the development of extrapulmonary acute lung injury. Shock. 2011;36:138–43.
Perl M, Chung CS, Perl U, Lomas-Neira J, de Paepe M, Cioffi WG, et al. Fas-induced pulmonary apoptosis and inflammation during indirect acute lung injury. Am J Respir Crit Care Med. 2007;176:591–601.
Perl M, Chung CS, Lomas-Neira J, Rachel TM, Biffl WL, Cioffi WG, et al. Silencing of Fas, but not caspase-8, in lung epithelial cells ameliorates pulmonary apoptosis, inflammation, and neutrophil influx after hemorrhagic shock and sepsis. Am J Pathol. 2005;167:1545–59.
Messer MP, Kellermann P, Weber SJ, Hohmann C, Denk S, Klohs B, et al. Silencing of fas, fas-associated via death domain, or caspase 3 differentially affects lung inflammation, apoptosis, and development of trauma-induced septic acute lung injury. Shock. 2013;39:19–27.
Matsuda N, Yamamoto S, Takano K, Kageyama S, Kurobe Y, Yoshihara Y, et al. Silencing of fas-associated death domain protects mice from septic lung inflammation and apoptosis. Am J Respir Crit Care Med. 2009;179:806–15.
Ehrhardt H, Pritzke T, Oak P, Kossert M, Biebach L, Förster K, et al. Absence of TNF-α enhances inflammatory response in the newborn lung undergoing mechanical ventilation. Am J Physiol Lung Cell Mol Physiol. 2016;310:L909–18.
Mao Q, Gundavarapu S, Patel C, Tsai A, Luks FI, De Paepe ME. The Fas system confers protection against alveolar disruption in hyperoxia-exposed newborn mice. Am J Respir Cell Mol Biol. 2008;39:717–29.
Guthmann F, Wissel H, Rüstow B. Early subcutaneous administration of etanercept (Enbrel) prevents from hyperoxia-induced lung injury. Exp Lung Res. 2009;35:770–80.
Kaya G, Saldir M, Polat A, Fidanci MK, Erdem A, Erdem G, et al. Evaluation of Etanercept Treatment in Newborn Rat Model with Hyperoxic Lung Injury. Fetal Pediatr Pathol. 2016;35:327–38.
Wolthuis EK, Vlaar AP, Choi G, Roelofs JJ, Haitsma JJ, van der Poll T, et al. Recombinant human soluble tumor necrosis factor-alpha receptor fusion protein partly attenuates ventilator-induced lung injury. Shock. 2009;31:262–6.
Pryhuber GS, O’Brien DP, Baggs R, Phipps R, Huyck H, Sanz I, et al. Ablation of tumor necrosis factor receptor type I (p55) alters oxygen-induced lung injury. Am J Physiol Lung Cell Mol Physiol. 2000;278:L1082–90.
Redente EF, Chakraborty S, Sajuthi S, Black BP, Edelman BL, Seibold MA, et al. Loss of Fas signaling in fibroblasts impairs homeostatic fibrosis resolution and promotes persistent pulmonary fibrosis. JCI Insight. 2020;6:e141618.
Hao Z, Hampel B, Yagita H, Rajewsky K. T cell-specific ablation of Fas leads to Fas ligand-mediated lymphocyte depletion and inflammatory pulmonary fibrosis. J Exp Med. 2004;199:1355–65.
Aoshiba K, Yasui S, Tamaoki J, Nagai A. The Fas/Fas-ligand system is not required for bleomycin-induced pulmonary fibrosis in mice. Am J Respir Crit Care Med. 2000;162:695–700.
Kuwano K, Hagimoto N, Kawasaki M, Yatomi T, Nakamura N, Nagata S, et al. Essential roles of the Fas-Fas ligand pathway in the development of pulmonary fibrosis. J Clin Investig. 1999;104:13–19.
Redente EF, Keith RC, Janssen W, Henson PM, Ortiz LA, Downey GP, et al. Tumor necrosis factor-α accelerates the resolution of established pulmonary fibrosis in mice by targeting profibrotic lung macrophages. Am J Respir Cell Mol Biol. 2014;50:825–37.
Oikonomou N, Harokopos V, Zalevsky J, Valavanis C, Kotanidou A, Szymkowski DE, et al. Soluble TNF mediates the transition from pulmonary inflammation to fibrosis. PLoS ONE. 2006;1:e108.
Kuroki M, Noguchi Y, Shimono M, Tomono K, Tashiro T, Obata Y, et al. Repression of bleomycin-induced pneumopathy by TNF. J Immunol. 2003;170:567–74.
Collison AM, Li J, de Siqueira AP, Lv X, Toop HD, Morris JC, et al. TRAIL signals through the ubiquitin ligase MID1 to promote pulmonary fibrosis. BMC Pulm Med. 2019;19:31.
McGrath EE, Lawrie A, Marriott HM, Mercer P, Cross SS, Arnold N, et al. Deficiency of tumour necrosis factor-related apoptosis-inducing ligand exacerbates lung injury and fibrosis. Thorax. 2012;67:796–803.
Malaviya R, Sunil VR, Venosa A, Verissimo VL, Cervelli JA, Vayas KN, et al. Attenuation of Nitrogen Mustard-Induced Pulmonary Injury and Fibrosis by Anti-Tumor Necrosis Factor-α Antibody. Toxicol Sci. 2015;148:71–88.
Tan J, Ni X. TNF-α antagonist may not be suitable for severe rituximab-induced interstitial lung disease. J Clin Pharm Ther. 2015;40:249–50.
Santos LD, Antunes KH, Muraro SP, de Souza GF, da Silva AG, Felipe JS, et al. TNF-mediated alveolar macrophage necroptosis drives disease pathogenesis during respiratory syncytial virus infection. The Eur Respir J. 2021;57:2003764.
Morris DR, Ansar M, Ivanciuc T, Qu Y, Casola A, Garofalo RP. Selective blockade of TNFR1 improves clinical disease and bronchoconstriction in experimental RSV infection. Viruses. 2020;12:1176.
Nguyen TH, Maltby S, Simpson JL, Eyers F, Baines KJ, Gibson PG, et al. TNF-α and macrophages are critical for respiratory syncytial virus-induced exacerbations in a mouse model of allergic airways disease. J Immunol. 2016;196:3547–58.
van den Berg E, van Woensel JB, Bos AP, Bem RA, Altemeier WA, Gill SE, et al. Role of the Fas/FasL system in a model of RSV infection in mechanically ventilated mice. Am J Physiol Lung Cell Mol Physiol. 2011;301:L451–460.
Lopez AD, Avasarala S, Grewal S, Murali AK, London L. Differential role of the Fas/Fas ligand apoptotic pathway in inflammation and lung fibrosis associated with reovirus 1/L-induced bronchiolitis obliterans organizing pneumonia and acute respiratory distress syndrome. J Immunol. 2009;183:8244–57.
Bem RA, Bos AP, Wösten-van Asperen RM, Bruijn M, Lutter R, Sprick MR, et al. Potential role of soluble TRAIL in epithelial injury in children with severe RSV infection. Am J Respir Cell Mol Biol. 2010;42:697–705.
Neuzil KM, Tang YW, Graham BS. Protective Role of TNF-alpha in respiratory syncytial virus infection in vitro and in vivo. Am J Med Sci. 1996;311:201–4.
Pant K, Chandrasekaran A, Chang CJ, Vageesh A, Popkov AJ, Weinberg JB. Effects of tumor necrosis factor on viral replication and pulmonary inflammation during acute mouse adenovirus type 1 respiratory infection. Virology. 2020;547:12–19.
Adkins LJ, Molloy CT, Weinberg JB. Fas activity mediates airway inflammation during mouse adenovirus type 1 respiratory infection. Virology. 2018;521:129–37.
Li XM, Chen X, Gu W, Guo YJ, Cheng Y, Peng J, et al. Impaired TNF/TNFR2 signaling enhances Th2 and Th17 polarization and aggravates allergic airway inflammation. Am J Physiol Lung Cell Mol Physiol. 2017;313:L592–l601.
Starkhammar M, Kumlien Georén S, Dahlén SE, Cardell LO, Adner M. TNFα-blockade stabilizes local airway hyperresponsiveness during TLR-induced exacerbations in murine model of asthma. Respir Res. 2015;16:129.
Faustino L, Fonseca DM, Florsheim EB, Resende RR, Lepique AP, Faquim-Mauro E, et al. Tumor necrosis factor-related apoptosis-inducing ligand mediates the resolution of allergic airway inflammation induced by chronic allergen inhalation. Mucosal Immunol. 2014;7:1199–208.
Yilmaz O, Karaman M, Bagriyanik HA, Firinci F, Kiray M, Turkeli A, et al. Comparison of TNF antagonism by etanercept and dexamethasone on airway epithelium and remodeling in an experimental model of asthma. Int Immunopharmacol. 2013;17:768–73.
Sharma SK, Almeida FA, Kierstein S, Hortobagyi L, Lin T, Larkin A, et al. Systemic FasL neutralization increases eosinophilic inflammation in a mouse model of asthma. Allergy. 2012;67:328–35.
Hwang SJ, Kim HS, Chung DH. Fas/Fas ligand-mediated apoptosis promotes hypersensitivity pneumonitis in mice by enhancing maturation of dendritic cells. Am J Respir Crit Care Med. 2010;181:1250–61.
Weckmann M, Collison A, Simpson JL, Kopp MV, Wark PA, Smyth MJ, et al. Critical link between TRAIL and CCL20 for the activation of TH2 cells and the expression of allergic airway disease. Nat Med. 2007;13:1308–15.
Chuang YH, Suen JL, Chiang BL. Fas-ligand-expressing adenovirus-transfected dendritic cells decrease allergen-specific T cells and airway inflammation in a murine model of asthma. J Mol Med. 2006;84:595–603.
Broide DH, Stachnick G, Castaneda D, Nayar J, Sriramarao P. Inhibition of eosinophilic inflammation in allergen-challenged TNF receptor p55/p75-and TNF receptor p55-deficient mice. Am J Respir Cell Mol Biol. 2001;24:304–11.
Whitehead GS, Thomas SY, Shalaby KH, Nakano K, Moran TP, Ward JM, et al. TNF is required for TLR ligand-mediated but not protease-mediated allergic airway inflammation. J Clin Investig. 2017;127:3313–26.
Maillet I, Schnyder-Candrian S, Couillin I, Quesniaux VF, Erard F, Moser R, et al. Allergic lung inflammation is mediated by soluble tumor necrosis factor (TNF) and attenuated by dominant-negative TNF biologics. Am J Respir Cell Mol Biol. 2011;45:731–9.
Choi IW, Sun K, Kim YS, Ko HM, Im SY, Kim JH, et al. TNF-alpha induces the late-phase airway hyperresponsiveness and airway inflammation through cytosolic phospholipase A(2) activation. J Allergy Clin Immunol. 2005;116:537–43.
Hildebrandt GC, Olkiewicz KM, Corrion L, Clouthier SG, Pierce EM, Liu C, et al. A role for TNF receptor type II in leukocyte infiltration into the lung during experimental idiopathic pneumonia syndrome. Biol Blood Marrow Transplant. 2008;14:385–96.
Wu Y, Shen Y, Zhang J, Wan C, Wang T, Xu D, et al. Increased serum TRAIL and DR5 levels correlated with lung function and inflammation in stable COPD patients. Int J Chronic Obstr Pulm Dis. 2015;10:2405–12.
Haw TJ, Starkey MR, Nair PM, Pavlidis S, Liu G, Nguyen DH, et al. A pathogenic role for tumor necrosis factor-related apoptosis-inducing ligand in chronic obstructive pulmonary disease. Mucosal Immunol. 2016;9:859–72.
Galluzzi L, Kepp O, Kroemer G. Mitochondrial regulation of cell death: a phylogenetically conserved control. Microb Cell. 2016;3:101–8.
Dadsena S, Zollo C, García-Sáez AJ. Mechanisms of mitochondrial cell death. Biochem Soc Trans. 2021;49:663–74.
Czabotar PE, Lessene G, Strasser A, Adams JM. Control of apoptosis by the BCL-2 protein family: implications for physiology and therapy. Nat Rev Mol Cell Biol. 2014;15:49–63.
Shamas-Din A, Kale J, Leber B, Andrews DW. Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb Perspect Biol. 2013;5:a008714.
Kalkavan H, Green DR. MOMP, cell suicide as a BCL-2 family business. Cell Death Differ. 2018;25:46–55.
Birkinshaw RW, Czabotar PE. The BCL-2 family of proteins and mitochondrial outer membrane permeabilisation. Semin Cell Dev Biol. 2017;72:152–62.
Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol. 2008;9:47–59.
Julien O, Wells JA. Caspases and their substrates. Cell Death Differ. 2017;24:1380–9.
Shalini S, Dorstyn L, Dawar S. Kumar S. Old, new and emerging functions of caspases. Cell Death Differ. 2015;22:526–39.
Green DR. Caspases and their substrates. Cold Spring Harb Perspect Biol. 2022;14:a041012.
Moldoveanu T, Czabotar PE. BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins. Cold Spring Harb Perspect Biol. 2020;12:a036319.
Llambi F, Wang YM, Victor B, Yang M, Schneider DM, Gingras S, et al. BOK is a non-canonical BCL-2 family effector of apoptosis regulated by ER-associated degradation. Cell. 2016;165:421–33.
Bleicken S, Landeta O, Landajuela A, Basañez G, García-Sáez AJ. Proapoptotic Bax and Bak proteins form stable protein-permeable pores of tunable size. J Biol Chem. 2013;288:33241–52.
Bleicken S, Wagner C, García-Sáez AJ. Mechanistic differences in the membrane activity of Bax and Bcl-xL correlate with their opposing roles in apoptosis. Biophys J. 2013;104:421–31.
Dewson G, Kratina T, Czabotar P, Day CL, Adams JM, Kluck RM. Bak activation for apoptosis involves oligomerization of dimers via their alpha6 helices. Mol Cell. 2009;36:696–703.
Dewson G, Kluck RM. Mechanisms by which Bak and Bax permeabilise mitochondria during apoptosis. J cell Sci. 2009;122:2801–8.
Verhagen AM, Ekert PG, Pakusch M, Silke J, Connolly LM, Reid GE, et al. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell. 2000;102:43–53.
Dorstyn L, Akey CW, Kumar S. New insights into apoptosome structure and function. Cell Death Differ. 2018;25:1194–208.
Shiozaki EN, Shi Y. Caspases, IAPs and Smac/DIABLO: mechanisms from structural biology. Trends Biochem Sci. 2004;29:486–94.
Huang DC, Strasser A. BH3-Only proteins-essential initiators of apoptotic cell death. Cell. 2000;103:839–42.
Kale J, Osterlund EJ, Andrews DW. BCL-2 family proteins: changing partners in the dance towards death. Cell Death Differ. 2018;25:65–80.
Giam M, Huang DC, Bouillet P. BH3-only proteins and their roles in programmed cell death. Oncogene. 2008;27:S128–136.
Gavathiotis E, Suzuki M, Davis ML, Pitter K, Bird GH, Katz SG, et al. BAX activation is initiated at a novel interaction site. Nature. 2008;455:1076–81.
Gavathiotis E, Reyna DE, Davis ML, Bird GH, Walensky LD. BH3-triggered structural reorganization drives the activation of proapoptotic BAX. Mol Cell. 2010;40:481–92.
Kim H, Tu HC, Ren D, Takeuchi O, Jeffers JR, Zambetti GP, et al. Stepwise activation of BAX and BAK by tBID, BIM, and PUMA initiates mitochondrial apoptosis. Mol Cell. 2009;36:487–99.
Wei MC, Zong WX, Cheng EH, Lindsten T, Panoutsakopoulou V, Ross AJ, et al. Proapoptotic BAX and BAK: a requisite gateway to mitochondrial dysfunction and death. Science. 2001;292:727–30.
Kim H, Rafiuddin-Shah M, Tu HC, Jeffers JR, Zambetti GP, Hsieh JJ, et al. Hierarchical regulation of mitochondrion-dependent apoptosis by BCL-2 subfamilies. Nat Cell Biol. 2006;8:1348–58.
Dai H, Smith A, Meng XW, Schneider PA, Pang YP, Kaufmann SH. Transient binding of an activator BH3 domain to the Bak BH3-binding groove initiates Bak oligomerization. J Cell Biol. 2011;194:39–48.
Chen HC, Kanai M, Inoue-Yamauchi A, Tu HC, Huang Y, Ren D, et al. An interconnected hierarchical model of cell death regulation by the BCL-2 family. Nat Cell Biol. 2015;17:1270–81.
O’Neill KL, Huang K, Zhang J, Chen Y, Luo X. Inactivation of prosurvival Bcl-2 proteins activates Bax/Bak through the outer mitochondrial membrane. Genes Dev. 2016;30:973–88.
Letai A, Bassik MC, Walensky LD, Sorcinelli MD, Weiler S, Korsmeyer SJ. Distinct BH3 domains either sensitize or activate mitochondrial apoptosis, serving as prototype cancer therapeutics. Cancer Cell. 2002;2:183–92.
Kuwana T, Bouchier-Hayes L, Chipuk JE, Bonzon C, Sullivan BA, Green DR, et al. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol Cell. 2005;17:525–35.
Chen L, Willis SN, Wei A, Smith BJ, Fletcher JI, Hinds MG, et al. Differential targeting of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic function. Mol Cell. 2005;17:393–403.
Jeng PS, Inoue-Yamauchi A, Hsieh JJ, Cheng EH. BH3-Dependent and Independent Activation of BAX and BAK in Mitochondrial. Apoptosis Curr Opin Physiol. 2018;3:71–81.
Vaseva AV, Moll UM. The mitochondrial p53 pathway. Biochim Biophys Acta. 2009;1787:414–20.
Mihara M, Erster S, Zaika A, Petrenko O, Chittenden T, Pancoska P, et al. p53 has a direct apoptogenic role at the mitochondria. Mol Cell. 2003;11:577–90.
Chen ZX, Pervaiz S. Involvement of cytochrome c oxidase subunits Va and Vb in the regulation of cancer cell metabolism by Bcl-2. Cell Death Differ. 2010;17:408–20.
Chong SJF, Iskandar K, Lai JXH, Qu J, Raman D, Valentin R, et al. Serine-70 phosphorylated Bcl-2 prevents oxidative stress-induced DNA damage by modulating the mitochondrial redox metabolism. Nucleic Acids Res. 2020;48:12727–45.
Clément MV, Hirpara JL, Pervaiz S. Decrease in intracellular superoxide sensitizes Bcl-2-overexpressing tumor cells to receptor and drug-induced apoptosis independent of the mitochondria. Cell Death Differ. 2003;10:1273–85.
Low IC, Loh T, Huang Y, Virshup DM, Pervaiz S. Ser70 phosphorylation of Bcl-2 by selective tyrosine nitration of PP2A-B56δ stabilizes its antiapoptotic activity. Blood. 2014;124:2223–34.
Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ. Bax-deficient mice with lymphoid hyperplasia and male germ cell death. Science. 1995;270:96–9.
Lindsten T, Ross AJ, King A, Zong WX, Rathmell JC, Shiels HA, et al. The combined functions of proapoptotic Bcl-2 family members bak and bax are essential for normal development of multiple tissues. Mol Cell. 2000;6:1389–99.
Ke F, Voss A, Kerr JB, O’Reilly LA, Tai L, Echeverry N, et al. BCL-2 family member BOK is widely expressed but its loss has only minimal impact in mice. Cell Death Differ. 2012;19:915–25.
Nguyen DH, Soygur B, Peng SP, Malki S, Hu G, Laird DJ. Apoptosis in the fetal testis eliminates developmentally defective germ cell clones. Nat Cell Biol. 2020;22:1423–35.
Russell LD, Chiarini-Garcia H, Korsmeyer SJ, Knudson CM. Bax-dependent spermatogonia apoptosis is required for testicular development and spermatogenesis. Biol Reprod. 2002;66:950–8.
Rodriguez I, Ody C, Araki K, Garcia I, Vassalli P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J. 1997;16:2262–70.
White FA, Keller-Peck CR, Knudson CM, Korsmeyer SJ, Snider WD. Widespread elimination of naturally occurring neuronal death in Bax-deficient mice. J Neurosci. 1998;18:1428–39.
Fan H, Favero M, Vogel MW. Elimination of Bax expression in mice increases cerebellar purkinje cell numbers but not the number of granule cells. J Comp Neurol. 2001;436:82–91.
Jung AR, Kim TW, Rhyu IJ, Kim H, Lee YD, Vinsant S, et al. Misplacement of Purkinje cells during postnatal development in Bax knock-out mice: a novel role for programmed cell death in the nervous system? J Neurosci. 2008;28:2941–8.
Sun W, Winseck A, Vinsant S, Park OH, Kim H, Oppenheim RW. Programmed cell death of adult-generated hippocampal neurons is mediated by the proapoptotic gene Bax. J Neurosci. 2004;24:11205–13.
Chang MY, Sun W, Ochiai W, Nakashima K, Kim SY, Park CH, et al. Bcl-XL/Bax proteins direct the fate of embryonic cortical precursor cells. Mol Cell Biol. 2007;27:4293–305.
Southwell DG, Paredes MF, Galvao RP, Jones DL, Froemke RC, Sebe JY, et al. Intrinsically determined cell death of developing cortical interneurons. Nature. 2012;491:109–13.
Jyotika J, McCutcheon J, Laroche J, Blaustein JD, Forger NG. Deletion of the Bax gene disrupts sexual behavior and modestly impairs motor function in mice. Dev Neurobiol. 2007;67:1511–9.
Luedke AC, Boucher PO, Niel L, Holmes MM. Altered anxiety and defensive behaviors in Bax knockout mice. Behav Brain Res. 2013;239:115–20.
Krahe TE, Medina AE, Lantz CL, Filgueiras CC. Hyperactivity and depression-like traits in Bax KO mice. Brain Res. 2015;1625:246–54.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
Ke F, Bouillet P, Kaufmann T, Strasser A, Kerr J, Voss AK. Consequences of the combined loss of BOK and BAK or BOK and BAX. Cell Death Dis. 2013;4:e650.
Arakawa S, Tsujioka M, Yoshida T, Tajima-Sakurai H, Nishida Y, Matsuoka Y, et al. Role of Atg5-dependent cell death in the embryonic development of Bax/Bak double-knockout mice. Cell Death Differ. 2017;24:1598–608.
Rybstein MD, Bravo-San Pedro JM, Kroemer G, Galluzzi L. The autophagic network and cancer. Nat Cell Biol. 2018;20:243–51.
Galluzzi L, Green DR. Autophagy-independent functions of the autophagy machinery. Cell. 2019;177:1682–99.
Miller DR, Cramer SD, Thorburn A. The interplay of autophagy and non-apoptotic cell death pathways. Int Rev Cell Mol Biol. 2020;352:159–87.
Fairlie WD, Tran S, Lee EF. Crosstalk between apoptosis and autophagy signaling pathways. Int Rev Cell Mol Biol. 2020;352:115–58.
Gu Z, Serradj N, Ueno M, Liang M, Li J, Baccei ML, et al. Skilled movements require non-apoptotic Bax/Bak pathway-mediated corticospinal circuit reorganization. Neuron. 2017;94:626–641.e624.
Hahn P, Lindsten T, Ying GS, Bennett J, Milam AH, Thompson CB, et al. Proapoptotic bcl-2 family members, Bax and Bak, are essential for developmental photoreceptor apoptosis. Investig Ophthalmol Vis Sci. 2003;44:3598–605.
Hahn P, Lindsten T, Tolentino M, Thompson CB, Bennett J, Dunaief JL. Persistent fetal ocular vasculature in mice deficient in bax and bak. Arch Ophthalmol. 2005;123:797–802.
Ke FS, Holloway S, Uren RT, Wong AW, Little MH, Kluck RM, et al. The BCL-2 family member BID plays a role during embryonic development in addition to its BH3-only protein function by acting in parallel to BAX, BAK and BOK. EMBO J. 2022;41:e110300.
Flores-Romero H, Hohorst L, John M, Albert MC, King LE, Beckmann L, et al. BCL-2-family protein tBID can act as a BAX-like effector of apoptosis. EMBO J. 2022;41:e108690.
Suzuki M, Youle RJ, Tjandra N. Structure of Bax: coregulation of dimer formation and intracellular localization. Cell. 2000;103:645–54.
McDonnell JM, Fushman D, Milliman CL, Korsmeyer SJ, Cowburn D. Solution structure of the proapoptotic molecule BID: a structural basis for apoptotic agonists and antagonists. Cell. 1999;96:625–34.
Chou JJ, Li H, Salvesen GS, Yuan J, Wagner G. Solution structure of BID, an intracellular amplifier of apoptotic signaling. Cell. 1999;96:615–24.
Takeuchi O, Fisher J, Suh H, Harada H, Malynn BA, Korsmeyer SJ. Essential role of BAX,BAK in B cell homeostasis and prevention of autoimmune disease. Proc Natl Acad Sci USA. 2005;102:11272–7.
Biswas S, Shi Q, Matise L, Cleveland S, Dave U, Zinkel S. A role for proapoptotic Bax and Bak in T-cell differentiation and transformation. Blood. 2010;116:5237–46.
Kodama T, Hikita H, Kawaguchi T, Shigekawa M, Shimizu S, Hayashi Y, et al. Mcl-1 and Bcl-xL regulate Bak/Bax-dependent apoptosis of the megakaryocytic lineage at multistages. Cell Death Differ. 2012;19:1856–69.
Pleines I, Lebois M, Gangatirkar P, Au AE, Lane RM, Henley KJ, et al. Intrinsic apoptosis circumvents the functional decline of circulating platelets but does not cause the storage lesion. Blood. 2018;132:197–209.
Ke F, Grabow S, Kelly GL, Lin A, O’Reilly LA, Strasser A. Impact of the combined loss of BOK, BAX and BAK on the hematopoietic system is slightly more severe than compound loss of BAX and BAK. Cell Death Dis. 2015;6:e1938.
Rathmell JC, Lindsten T, Zong WX, Cinalli RM, Thompson CB. Deficiency in Bak and Bax perturbs thymic selection and lymphoid homeostasis. Nat Immunol. 2002;3:932–9.
Jones RG, Bui T, White C, Madesh M, Krawczyk CM, Lindsten T, et al. The proapoptotic factors Bax and Bak regulate T Cell proliferation through control of endoplasmic reticulum Ca(2+) homeostasis. Immunity. 2007;27:268–80.
Watson EC, Koenig MN, Grant ZL, Whitehead L, Trounson E, Dewson G, et al. Apoptosis regulates endothelial cell number and capillary vessel diameter but not vessel regression during retinal angiogenesis. Devlopment. 2016;143:2973–82.
Wang YM, Zhang GY, Wang Y, Hu M, Zhou JJ, Sawyer A, et al. Exacerbation of spontaneous autoimmune nephritis following regulatory T cell depletion in B cell lymphoma 2-interacting mediator knock-out mice. Clin Exp Immunol. 2017;188:195–207.
Puthalakath H, O’Reilly LA, Gunn P, Lee L, Kelly PN, Huntington ND, et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell. 2007;129:1337–49.
Hutcheson J, Perlman H. Loss of Bim results in abnormal accumulation of mature CD4-CD8-CD44-CD25- thymocytes. Immunobiology. 2007;212:629–36.
Chougnet CA, Tripathi P, Lages CS, Raynor J, Sholl A, Fink P, et al. A major role for Bim in regulatory T cell homeostasis. J Immunol. 2011;186:156–63.
Bouillet P, Purton JF, Godfrey DI, Zhang LC, Coultas L, Puthalakath H, et al. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature. 2002;415:922–6.
Enders A, Bouillet P, Puthalakath H, Xu Y, Tarlinton DM, Strasser A. Loss of the pro-apoptotic BH3-only Bcl-2 family member Bim inhibits BCR stimulation-induced apoptosis and deletion of autoreactive B cells. J Exp Med. 2003;198:1119–26.
Zhan Y, Zhang Y, Gray D, Carrington EM, Bouillet P, Ko HJ, et al. Defects in the Bcl-2-regulated apoptotic pathway lead to preferential increase of CD25 low Foxp3+ anergic CD4+ T cells. J Immunol. 2011;187:1566–77.
Pellegrini M, Belz G, Bouillet P, Strasser A. Shutdown of an acute T cell immune response to viral infection is mediated by the proapoptotic Bcl-2 homology 3-only protein Bim. Proc Natl Acad Sci USA. 2003;100:14175–80.
Hildeman DA, Zhu Y, Mitchell TC, Bouillet P, Strasser A, Kappler J, et al. Activated T cell death in vivo mediated by proapoptotic bcl-2 family member bim. Immunity. 2002;16:759–67.
Fischer SF, Bouillet P, O’Donnell K, Light A, Tarlinton DM, Strasser A. Proapoptotic BH3-only protein Bim is essential for developmentally programmed death of germinal center-derived memory B cells and antibody-forming cells. Blood. 2007;110:3978–84.
Sugimoto-Ishige A, Harada M, Tanaka M, Terooatea T, Adachi Y, Takahashi Y, et al. Bim establishes the B-cell repertoire from early to late in the immune response. Int Immunol. 2021;33:79–90.
Oliver PM, Wang M, Zhu Y, White J, Kappler J, Marrack P. Loss of Bim allows precursor B cell survival but not precursor B cell differentiation in the absence of interleukin 7. J Exp Med. 2004;200:1179–87.
Villunger A, Scott C, Bouillet P, Strasser A. Essential role for the BH3-only protein Bim but redundant roles for Bax, Bcl-2, and Bcl-w in the control of granulocyte survival. Blood. 2003;101:2393–400.
Mailleux AA, Overholtzer M, Schmelzle T, Bouillet P, Strasser A, Brugge JS. BIM regulates apoptosis during mammary ductal morphogenesis, and its absence reveals alternative cell death mechanisms. Dev Cell. 2007;12:221–34.
Schuler F, Baumgartner F, Klepsch V, Chamson M, Müller-Holzner E, Watson CJ, et al. The BH3-only protein BIM contributes to late-stage involution in the mouse mammary gland. Cell Death Differ. 2016;23:41–51.
Ohgushi M, Kuroki S, Fukamachi H, O’Reilly LA, Kuida K, Strasser A, et al. Transforming growth factor beta-dependent sequential activation of Smad, Bim, and caspase-9 mediates physiological apoptosis in gastric epithelial cells. Mol Cell Biol. 2005;25:10017–28.
Doonan F, Donovan M, Gomez-Vicente V, Bouillet P, Cotter TG. Bim expression indicates the pathway to retinal cell death in development and degeneration. J Neurosci. 2007;27:10887–94.
Wali JA, Galic S, Tan CY, Gurzov EN, Frazier AE, Connor T, et al. Loss of BIM increases mitochondrial oxygen consumption and lipid oxidation, reduces adiposity and improves insulin sensitivity in mice. Cell Death Differ. 2018;25:217–25.
Hutcheson J, Scatizzi JC, Bickel E, Brown NJ, Bouillet P, Strasser A, et al. Combined loss of proapoptotic genes Bak or Bax with Bim synergizes to cause defects in hematopoiesis and in thymocyte apoptosis. J Exp Med. 2005;201:1949–60.
Liu R, King A, Bouillet P, Tarlinton DM, Strasser A, Heierhorst J, Proapoptotic BIM. Impacts B lymphoid homeostasis by limiting the survival of mature B cells in a cell-autonomous manner. Front Immunol. 2018;9:592.
Herold MJ, Stuchbery R, Mérino D, Willson T, Strasser A, Hildeman D, et al. Impact of conditional deletion of the pro-apoptotic BCL-2 family member BIM in mice. Cell Death Dis. 2014;5:e1446.
Huntington ND, Labi V, Cumano A, Vieira P, Strasser A, Villunger A, et al. Loss of the pro-apoptotic BH3-only Bcl-2 family member Bim sustains B lymphopoiesis in the absence of IL-7. Int Immunol. 2009;21:715–25.
Ludwig LM, Roach LE, Katz SG, LaBelle JL. Loss of BIM in T cells results in BCL-2 family BH3-member compensation but incomplete cell death sensitivity normalization. Apoptosis. 2020;25:247–60.
Bunk EC, König HG, Bernas T, Engel T, Henshall DC, Kirby BP, et al. BH3-only proteins BIM and PUMA in the regulation of survival and neuronal differentiation of newly generated cells in the adult mouse hippocampus. Cell Death Dis. 2010;1:e15.
Tsai F, Homan PJ, Agrawal H, Misharin AV, Abdala-Valencia H, Haines GK 3rd, et al. Bim suppresses the development of SLE by limiting myeloid inflammatory responses. J Exp Med. 2017;214:3753–73.
Leonard JR, D’Sa C, Cahn BR, Korsmeyer SJ, Roth KA. Bid regulation of neuronal apoptosis. Brain Res Dev Brain Res. 2001;128:187–90.
Jeffers JR, Parganas E, Lee Y, Yang C, Wang J, Brennan J, et al. Puma is an essential mediator of p53-dependent and -independent apoptotic pathways. Cancer Cell. 2003;4:321–8.
McKenzie MD, Carrington EM, Kaufmann T, Strasser A, Huang DC, Kay TW, et al. Proapoptotic BH3-only protein Bid is essential for death receptor-induced apoptosis of pancreatic beta-cells. Diabetes. 2008;57:1284–92.
Jost PJ, Grabow S, Gray D, McKenzie MD, Nachbur U, Huang DC, et al. XIAP discriminates between type I and type II FAS-induced apoptosis. Nature. 2009;460:1035–9.
Zinkel SS, Ong CC, Ferguson DO, Iwasaki H, Akashi K, Bronson RT, et al. Proapoptotic BID is required for myeloid homeostasis and tumor suppression. Genes Dev. 2003;17:229–39.
Salisbury-Ruf CT, Bertram CC, Vergeade A, Lark DS, Shi Q, Heberling ML, et al. Bid maintains mitochondrial cristae structure and function and protects against cardiac disease in an integrative genomics study. eLife. 2018;7:e40907.
Tischner D, Gaggl I, Peschel I, Kaufmann M, Tuzlak S, Drach M, et al. Defective cell death signalling along the Bcl-2 regulated apoptosis pathway compromises Treg cell development and limits their functionality in mice. J Autoimmun. 2012;38:59–69.
Myers M, Morgan FH, Liew SH, Zerafa N, Gamage TU, Sarraj M, et al. PUMA regulates germ cell loss and primordial follicle endowment in mice. Reproduction. 2014;148:211–9.
Erlacher M, Michalak EM, Kelly PN, Labi V, Niederegger H, Coultas L, et al. BH3-only proteins Puma and Bim are rate-limiting for gamma-radiation- and glucocorticoid-induced apoptosis of lymphoid cells in vivo. Blood. 2005;106:4131–8.
Wang J, Thomas HR, Li Z, Yeo NCF, Scott HE, Dang N, et al. Puma, noxa, p53, and p63 differentially mediate stress pathway induced apoptosis. Cell Death Dis. 2021;12:659.
Kerr JB, Hutt KJ, Michalak EM, Cook M, Vandenberg CJ, Liew SH, et al. DNA damage-induced primordial follicle oocyte apoptosis and loss of fertility require TAp63-mediated induction of Puma and Noxa. Mol Cell. 2012;48:343–52.
Naik E, Michalak EM, Villunger A, Adams JM, Strasser A. Ultraviolet radiation triggers apoptosis of fibroblasts and skin keratinocytes mainly via the BH3-only protein Noxa. J Cell Biol. 2007;176:415–24.
Wensveen FM, Geest CR, Libregts S, Derks IAM, Ekert PG, Labi V, et al. BH3-only protein Noxa contributes to apoptotic control of stress-erythropoiesis. Apoptosis. 2013;18:1306–18.
Clybouw C, Fischer S, Auffredou MT, Hugues P, Alexia C, Bouillet P, et al. Regulation of memory B-cell survival by the BH3-only protein Puma. Blood. 2011;118:4120–8.
Wensveen FM, Derks IA, van Gisbergen KP, de Bruin AM, Meijers JC, Yigittop H, et al. BH3-only protein Noxa regulates apoptosis in activated B cells and controls high-affinity antibody formation. Blood. 2012;119:1440–9.
Michalak EM, Villunger A, Adams JM, Strasser A. In several cell types tumour suppressor p53 induces apoptosis largely via Puma but Noxa can contribute. Cell Death Differ. 2008;15:1019–29.
Erlacher M, Labi V, Manzl C, Böck G, Tzankov A, Häcker G, et al. Puma cooperates with Bim, the rate-limiting BH3-only protein in cell death during lymphocyte development, in apoptosis induction. J Exp Med. 2006;203:2939–51.
Gray DH, Kupresanin F, Berzins SP, Herold MJ, O’Reilly LA, Bouillet P, et al. The BH3-only proteins Bim and Puma cooperate to impose deletional tolerance of organ-specific antigens. Immunity. 2012;37:451–62.
Happo L, Cragg MS, Phipson B, Haga JM, Jansen ES, Herold MJ, et al. Maximal killing of lymphoma cells by DNA damage-inducing therapy requires not only the p53 targets Puma and Noxa, but also Bim. Blood. 2010;116:5256–67.
Ren D, Tu HC, Kim H, Wang GX, Bean GR, Takeuchi O, et al. BID, BIM, and PUMA are essential for activation of the BAX- and BAK-dependent cell death program. Science. 2010;330:1390–3.
Ranger AM, Zha J, Harada H, Datta SR, Danial NN, Gilmore AP, et al. Bad-deficient mice develop diffuse large B cell lymphoma. Proc Natl Acad Sci USA. 2003;100:9324–9.
Labi V, Erlacher M, Kiessling S, Manzl C, Frenzel A, O’Reilly L, et al. Loss of the BH3-only protein Bmf impairs B cell homeostasis and accelerates gamma irradiation-induced thymic lymphoma development. J Exp Med. 2008;205:641–55.
Coultas L, Bouillet P, Stanley EG, Brodnicki TC, Adams JM, Strasser A. Proapoptotic BH3-only Bcl-2 family member Bik/Blk/Nbk is expressed in hemopoietic and endothelial cells but is redundant for their programmed death. Mol Cell Biol. 2004;24:1570–81.
Kelly PN, White MJ, Goschnick MW, Fairfax KA, Tarlinton DM, Kinkel SA, et al. Individual and overlapping roles of BH3-only proteins Bim and Bad in apoptosis of lymphocytes and platelets and in suppression of thymic lymphoma development. Cell Death Differ. 2010;17:1655–64.
Hübner A, Cavanagh-Kyros J, Rincon M, Flavell RA, Davis RJ. Functional cooperation of the proapoptotic Bcl2 family proteins Bmf and Bim in vivo. Mol Cell Biol. 2010;30:98–105.
Vaithiyanathan K, Liew SH, Zerafa N, Gamage T, Cook M, O’Reilly LA, et al. BCL2-modifying factor promotes germ cell loss during murine oogenesis. Reproduction. 2016;151:553–62.
Liew SH, Vaithiyanathan K, Cook M, Bouillet P, Scott CL, Kerr JB, et al. Loss of the proapoptotic BH3-only protein BCL-2 modifying factor prolongs the fertile life span in female mice. Biol Reprod. 2014;90:77.
Baumgartner F, Woess C, Pedit V, Tzankov A, Labi V, Villunger A. Minor cell-death defects but reduced tumor latency in mice lacking the BH3-only proteins Bad and Bmf. Oncogene. 2013;32:621–30.
Coultas L, Bouillet P, Loveland KL, Meachem S, Perlman H, Adams JM, et al. Concomitant loss of proapoptotic BH3-only Bcl-2 antagonists Bik and Bim arrests spermatogenesis. EMBO J. 2005;24:3963–73.
Labi V, Woess C, Tuzlak S, Erlacher M, Bouillet P, Strasser A, et al. Deregulated cell death and lymphocyte homeostasis cause premature lethality in mice lacking the BH3-only proteins Bim and Bmf. Blood. 2014;123:2652–62.
Woess C, Tuzlak S, Labi V, Drach M, Bertele D, Schneider P, et al. Combined loss of the BH3-only proteins Bim and Bmf restores B-cell development and function in TACI-Ig transgenic mice. Cell Death Differ. 2015;22:1477–88.
Rinkenberger JL, Horning S, Klocke B, Roth K, Korsmeyer SJ. Mcl-1 deficiency results in peri-implantation embryonic lethality. Genes Dev. 2000;14:23–27.
Kuida K, Haydar TF, Kuan CY, Gu Y, Taya C, Karasuyama H, et al. Reduced apoptosis and cytochrome c-mediated caspase activation in mice lacking caspase 9. Cell. 1998;94:325–37.
Motoyama N, Wang F, Roth KA, Sawa H, Nakayama K, Nakayama K, et al. Massive cell death of immature hematopoietic cells and neurons in Bcl-x-deficient mice. Science. 1995;267:1506–10.
Zaidi AU, D’Sa-Eipper C, Brenner J, Kuida K, Zheng TS, Flavell RA, et al. Bcl-X(L)-caspase-9 interactions in the developing nervous system: evidence for multiple death pathways. J Neurosci. 2001;21:169–75.
Shindler KS, Latham CB, Roth KA. Bax deficiency prevents the increased cell death of immature neurons in bcl-x-deficient mice. J Neurosci. 1997;17:3112–9.
Akhtar RS, Klocke BJ, Strasser A, Roth KA. Loss of BH3-only protein Bim inhibits apoptosis of hemopoietic cells in the fetal liver and male germ cells but not neuronal cells in bcl-x-deficient mice. J Histochem Cytochem. 2008;56:921–7.
Bouillet P, Cory S, Zhang LC, Strasser A, Adams JM. Degenerative disorders caused by Bcl-2 deficiency prevented by loss of its BH3-only antagonist Bim. Dev Cell. 2001;1:645–53.
Veis DJ, Sorenson CM, Shutter JR, Korsmeyer SJ. Bcl-2-deficient mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell. 1993;75:229–40.
Nakayama K, Nakayama K, Negishi I, Kuida K, Shinkai Y, Louie MC, et al. Disappearance of the lymphoid system in Bcl-2 homozygous mutant chimeric mice. Science. 1993;261:1584–8.
Kamada S, Shimono A, Shinto Y, Tsujimura T, Takahashi T, Noda T, et al. bcl-2 deficiency in mice leads to pleiotropic abnormalities: accelerated lymphoid cell death in thymus and spleen, polycystic kidney, hair hypopigmentation, and distorted small intestine. Cancer Res. 1995;55:354–9.
Michaelidis TM, Sendtner M, Cooper JD, Airaksinen MS, Holtmann B, Meyer M, et al. Inactivation of bcl-2 results in progressive degeneration of motoneurons, sympathetic and sensory neurons during early postnatal development. Neuron. 1996;17:75–89.
Manzl C, Baumgartner F, Peintner L, Schuler F, Villunger A. Possible pitfalls investigating cell death responses in genetically engineered mouse models and derived cell lines. Methods. 2013;61:130–7.
Carpinelli MR, Wise AK, Arhatari BD, Bouillet P, Manji SS, Manning MG, et al. Anti-apoptotic gene Bcl2 is required for stapes development and hearing. Cell Death Dis. 2012;3:e362.
Hamasaki A, Sendo F, Nakayama K, Ishida N, Negishi I, Nakayama K, et al. Accelerated neutrophil apoptosis in mice lacking A1-a, a subtype of the bcl-2-related A1 gene. J Exp Med. 1998;188:1985–92.
Xiang Z, Ahmed AA, Möller C, Nakayama K, Hatakeyama S, Nilsson G. Essential role of the prosurvival bcl-2 homologue A1 in mast cell survival after allergic activation. J Exp Med. 2001;194:1561–9.
Schenk RL, Tuzlak S, Carrington EM, Zhan Y, Heinzel S, Teh CE, et al. Characterisation of mice lacking all functional isoforms of the pro-survival BCL-2 family member A1 reveals minor defects in the haematopoietic compartment. Cell Death Differ. 2017;24:534–45.
Tuzlak S, Schenk RL, Vasanthakumar A, Preston SP, Haschka MD, Zotos D, et al. The BCL-2 pro-survival protein A1 is dispensable for T cell homeostasis on viral infection. Cell Death Differ. 2017;24:523–33.
Print CG, Loveland KL, Gibson L, Meehan T, Stylianou A, Wreford N, et al. Apoptosis regulator bcl-w is essential for spermatogenesis but appears otherwise redundant. Proc Natl Acad Sci USA. 1998;95:12424–31.
Ross AJ, Waymire KG, Moss JE, Parlow AF, Skinner MK, Russell LD, et al. Testicular degeneration in Bclw-deficient mice. Nat Genet. 1998;18:251–6.
Russell LD, Warren J, Debeljuk L, Richardson LL, Mahar PL, Waymire KG, et al. Spermatogenesis in Bclw-deficient mice. Biol Reprod. 2001;65:318–32.
Brinkmann K, Grabow S, Hyland CD, Teh CE, Alexander WS, Herold MJ, et al. The combination of reduced MCL-1 and standard chemotherapeutics is tolerable in mice. Cell Death Differ. 2017;24:2032–43.
Delbridge AR, Opferman JT, Grabow S, Strasser A. Antagonism between MCL-1 and PUMA governs stem/progenitor cell survival during hematopoietic recovery from stress. Blood. 2015;125:3273–80.
Kasai S, Chuma S, Motoyama N, Nakatsuji N. Haploinsufficiency of Bcl-x leads to male-specific defects in fetal germ cells: differential regulation of germ cell apoptosis between the sexes. Dev Biol. 2003;264:202–16.
Mason KD, Carpinelli MR, Fletcher JI, Collinge JE, Hilton AA, Ellis S, et al. Programmed anuclear cell death delimits platelet life span. Cell. 2007;128:1173–86.
Schenk RL, Gangoda L, Lawlor KE, O’Reilly LA, Strasser A, Herold MJ. The pro-survival Bcl-2 family member A1 delays spontaneous and FAS ligand-induced apoptosis of activated neutrophils. Cell Death Dis. 2020;11:474.
Grabow S, Kueh AJ, Ke F, Vanyai HK, Sheikh BN, Dengler MA, et al. Subtle Changes in the Levels of BCL-2 proteins cause severe craniofacial abnormalities. Cell Rep. 2018;24:3285–3295.e3284.
Ke F, Lancaster GI, Grabow S, Murphy AJ, Strasser A. Combined reduction in the expression of MCL-1 and BCL-2 reduces organismal size in mice. Cell Death Dis. 2020;11:185.
Opferman JT, Iwasaki H, Ong CC, Suh H, Mizuno S, Akashi K, et al. Obligate role of anti-apoptotic MCL-1 in the survival of hematopoietic stem cells. Science. 2005;307:1101–4.
Opferman JT, Letai A, Beard C, Sorcinelli MD, Ong CC, Korsmeyer SJ. Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature. 2003;426:671–6.
Pierson W, Cauwe B, Policheni A, Schlenner SM, Franckaert D, Berges J, et al. Antiapoptotic Mcl-1 is critical for the survival and niche-filling capacity of Foxp3+ regulatory T cells. Nat Immunol. 2013;14:959–65.
Vikstrom I, Carotta S, Lüthje K, Peperzak V, Jost PJ, Glaser S, et al. Mcl-1 is essential for germinal center formation and B cell memory. Science. 2010;330:1095–9.
Tripathi P, Koss B, Opferman JT, Hildeman DA. Mcl-1 antagonizes Bax/Bak to promote effector CD4(+) and CD8(+) T-cell responses. Cell Death Differ. 2013;20:998–1007.
Dunkle A, Dzhagalov I, He YW. Mcl-1 promotes survival of thymocytes by inhibition of Bak in a pathway separate from Bcl-2. Cell Death Differ. 2010;17:994–1002.
Sathe P, Delconte RB, Souza-Fonseca-Guimaraes F, Seillet C, Chopin M, Vandenberg CJ, et al. Innate immunodeficiency following genetic ablation of Mcl1 in natural killer cells. Nat Commun. 2014;5:4539.
Dzhagalov I, St John A, He YW. The antiapoptotic protein Mcl-1 is essential for the survival of neutrophils but not macrophages. Blood. 2007;109:1620–6.
Steimer DA, Boyd K, Takeuchi O, Fisher JK, Zambetti GP, Opferman JT. Selective roles for antiapoptotic MCL-1 during granulocyte development and macrophage effector function. Blood. 2009;113:2805–15.
Lilla JN, Chen CC, Mukai K, BenBarak MJ, Franco CB, Kalesnikoff J, et al. Reduced mast cell and basophil numbers and function in Cpa3-Cre; Mcl-1fl/fl mice. Blood. 2011;118:6930–8.
Slomp A, Peperzak V. Role and regulation of pro-survival BCL-2 Proteins in Multiple Myeloma. Front Oncol. 2018;8:533.
Peperzak V, Vikström I, Walker J, Glaser SP, LePage M, Coquery CM, et al. Mcl-1 is essential for the survival of plasma cells. Nat Immunol. 2013;14:290–7.
Carrington EM, Zhan Y, Brady JL, Zhang JG, Sutherland RM, Anstee NS, et al. Anti-apoptotic proteins BCL-2, MCL-1 and A1 summate collectively to maintain survival of immune cell populations both in vitro and in vivo. Cell Death Differ. 2017;24:878–88.
Dzhagalov I, Dunkle A, He YW. The anti-apoptotic Bcl-2 family member Mcl-1 promotes T lymphocyte survival at multiple stages. J Immunol. 2008;181:521–8.
Malin S, McManus S, Cobaleda C, Novatchkova M, Delogu A, Bouillet P, et al. Role of STAT5 in controlling cell survival and immunoglobulin gene recombination during pro-B cell development. Nat Immunol. 2010;11:171–9.
Debrincat MA, Josefsson EC, James C, Henley KJ, Ellis S, Lebois M, et al. Mcl-1 and Bcl-x(L) coordinately regulate megakaryocyte survival. Blood. 2012;119:5850–8.
Josefsson EC, James C, Henley KJ, Debrincat MA, Rogers KL, Dowling MR, et al. Megakaryocytes possess a functional intrinsic apoptosis pathway that must be restrained to survive and produce platelets. J Exp Med. 2011;208:2017–31.
Wagner KU, Claudio E, Rucker EB 3rd, Riedlinger G, Broussard C, Schwartzberg PL, et al. Conditional deletion of the Bcl-x gene from erythroid cells results in hemolytic anemia and profound splenomegaly. Development. 2000;127:4949–58.
Vikström IB, Slomp A, Carrington EM, Moesbergen LM, Chang C, Kelly GL, et al. MCL-1 is required throughout B-cell development and its loss sensitizes specific B-cell subsets to inhibition of BCL-2 or BCL-XL. Cell Death Dis. 2016;7:e2345.
Sochalska M, Ottina E, Tuzlak S, Herzog S, Herold M, Villunger A. Conditional knockdown of BCL2A1 reveals rate-limiting roles in BCR-dependent B-cell survival. Cell Death Differ. 2016;23:628–39.
Debrincat MA, Pleines I, Lebois M, Lane RM, Holmes ML, Corbin J, et al. BCL-2 is dispensable for thrombopoiesis and platelet survival. Cell Death Dis. 2015;6:e1721.
Ma A, Pena JC, Chang B, Margosian E, Davidson L, Alt FW, et al. Bclx regulates the survival of double-positive thymocytes. Proc Natl Acad Sci USA. 1995;92:4763–7.
Matsuzaki Y, Nakayama K, Nakayama K, Tomita T, Isoda M, Loh DY, et al. Role of bcl-2 in the development of lymphoid cells from the hematopoietic stem cell. Blood. 1997;89:853–62.
Geueke A, Mantellato G, Kuester F, Schettina P, Nelles M, Seeger JM, et al. The anti-apoptotic Bcl-2 protein regulates hair follicle stem cell function. EMBO Rep. 2021;22:e52301.
Thomas RL, Gustafsson AB. MCL1 is critical for mitochondrial function and autophagy in the heart. Autophagy. 2013;9:1902–3.
Wang X, Bathina M, Lynch J, Koss B, Calabrese C, Frase S, et al. Deletion of MCL-1 causes lethal cardiac failure and mitochondrial dysfunction. Genes Dev. 2013;27:1351–64.
Arbour N, Vanderluit JL, Le Grand JN, Jahani-Asl A, Ruzhynsky VA, Cheung EC, et al. Mcl-1 is a key regulator of apoptosis during CNS development and after DNA damage. J Neurosci. 2008;28:6068–78.
Germain M, Nguyen AP, Le Grand JN, Arbour N, Vanderluit JL, Park DS, et al. MCL-1 is a stress sensor that regulates autophagy in a developmentally regulated manner. EMBO J. 2011;30:395–407.
Malone CD, Hasan SM, Roome RB, Xiong J, Furlong M, Opferman JT, et al. Mcl-1 regulates the survival of adult neural precursor cells. Mol Cell Neurosci. 2012;49:439–47.
Nakamura A, Swahari V, Plestant C, Smith I, McCoy E, Smith S, et al. Bcl-xL is essential for the survival and function of differentiated neurons in the cortex that control complex behaviors. J Neurosci. 2016;36:5448–61.
Savitt JM, Jang SS, Mu W, Dawson VL, Dawson TM. Bcl-x is required for proper development of the mouse substantia nigra. J Neurosci. 2005;25:6721–8.
Fogarty LC, Song B, Suppiah Y, Hasan SMM, Martin HC, Hogan SE, et al. Bcl-xL dependency coincides with the onset of neurogenesis in the developing mammalian spinal cord. Mol Cell Neurosci. 2016;77:34–46.
Fogarty LC, Flemmer RT, Geizer BA, Licursi M, Karunanithy A, Opferman JT, et al. Mcl-1 and Bcl-xL are essential for survival of the developing nervous system. Cell Death Differ. 2019;26:1501–15.
Veleta KA, Cleveland AH, Babcock BR, He YW, Hwang D, Sokolsky-Papkov M, et al. Antiapoptotic Bcl-2 family proteins BCL-xL and MCL-1 integrate neural progenitor survival and proliferation during postnatal cerebellar neurogenesis. Cell Death Differ. 2021;28:1579–92.
Hikita H, Takehara T, Shimizu S, Kodama T, Li W, Miyagi T, et al. Mcl-1 and Bcl-xL cooperatively maintain integrity of hepatocytes in developing and adult murine liver. Hepatology. 2009;50:1217–26.
Takehara T, Tatsumi T, Suzuki T, Rucker EB 3rd, Hennighausen L, Jinushi M, et al. Hepatocyte-specific disruption of Bcl-xL leads to continuous hepatocyte apoptosis and liver fibrotic responses. Gastroenterology. 2004;127:1189–97.
Vick B, Weber A, Urbanik T, Maass T, Teufel A, Krammer PH, et al. Knockout of myeloid cell leukemia-1 induces liver damage and increases apoptosis susceptibility of murine hepatocytes. Hepatology. 2009;49:627–36.
Watson EC, Whitehead L, Adams RH, Dewson G, Coultas L. Endothelial cell survival during angiogenesis requires the pro-survival protein MCL1. Cell Death Differ. 2016;23:1371–9.
Jain R, Sheridan JM, Policheni A, Heinlein M, Gandolfo LC, Dewson G, et al. A critical epithelial survival axis regulated by MCL-1 maintains thymic function in mice. Blood. 2017;130:2504–15.
Healy ME, Boege Y, Hodder MC, Böhm F, Malehmir M, Scherr AL, et al. MCL1 is required for maintenance of intestinal homeostasis and prevention of carcinogenesis in mice. Gastroenterology. 2020;159:183–99.
Walton KD, Wagner KU, Rucker EB 3rd, Shillingford JM, Miyoshi K, Hennighausen L. Conditional deletion of the bcl-x gene from mouse mammary epithelium results in accelerated apoptosis during involution but does not compromise cell function during lactation. Mech Dev. 2001;109:281–93.
Fu NY, Rios AC, Pal B, Soetanto R, Lun AT, Liu K, et al. EGF-mediated induction of Mcl-1 at the switch to lactation is essential for alveolar cell survival. Nat Cell Biol. 2015;17:365–75.
Staversky RJ, Vitiello PF, Yee M, Callahan LM, Dean DA, O’Reilly MA. Epithelial ablation of Bcl-XL increases sensitivity to oxygen without disrupting lung development. Am J Respir Cell Mol Biol. 2010;43:376–85.
Turnis ME, Kaminska E, Smith KH, Kartchner BJ, Vogel P, Laxton JD, et al. Requirement for antiapoptotic MCL-1 during early erythropoiesis. Blood. 2021;137:1945–58.
Teh CE, Robbins AK, Henstridge DC, Dewson G, Diepstraten ST, Kelly G, et al. MCL-1 is essential for survival but dispensable for metabolic fitness of FOXP3(+) regulatory T cells. Cell Death Differ. 2020;27:3374–85.
Hikita H, Takehara T, Kodama T, Shimizu S, Hosui A, Miyagi T, et al. BH3-only protein bid participates in the Bcl-2 network in healthy liver cells. Hepatology. 2009;50:1972–80.
Kodama T, Hikita H, Kawaguchi T, Saito Y, Tanaka S, Shigekawa M, et al. The Bcl-2 homology domain 3 (BH3)-only proteins Bim and bid are functionally active and restrained by anti-apoptotic Bcl-2 family proteins in healthy liver. J Biol Chem. 2013;288:30009–18.
Weber A, Boger R, Vick B, Urbanik T, Haybaeck J, Zoller S, et al. Hepatocyte-specific deletion of the antiapoptotic protein myeloid cell leukemia-1 triggers proliferation and hepatocarcinogenesis in mice. Hepatology. 2010;51:1226–36.
Coussens LM, Zitvogel L, Palucka AK. Neutralizing tumor-promoting chronic inflammation: a magic bullet? Science. 2013;339:286–91.
Cecconi F, Alvarez-Bolado G, Meyer BI, Roth KA, Gruss P. Apaf1 (CED-4 homolog) regulates programmed cell death in mammalian development. Cell. 1998;94:727–37.
Honarpour N, Du C, Richardson JA, Hammer RE, Wang X, Herz J. Adult Apaf-1-deficient mice exhibit male infertility. Dev Biol. 2000;218:248–58.
Yoshida H, Kong YY, Yoshida R, Elia AJ, Hakem A, Hakem R, et al. Apaf1 is required for mitochondrial pathways of apoptosis and brain development. Cell. 1998;94:739–50.
Cecconi F, Roth KA, Dolgov O, Munarriz E, Anokhin K, Gruss P, et al. Apaf1-dependent programmed cell death is required for inner ear morphogenesis and growth. Development. 2004;131:2125–35.
Long AB, Kaiser WJ, Mocarski ES, Caspary T. Apaf1 apoptotic function critically limits Sonic hedgehog signaling during craniofacial development. Cell Death Differ. 2013;20:1510–20.
Ohsawa S, Hamada S, Kuida K, Yoshida H, Igaki T, Miura M. Maturation of the olfactory sensory neurons by Apaf-1/caspase-9-mediated caspase activity. Proc Natl Acad Sci USA. 2010;107:13366–71.
Hakem R, Hakem A, Duncan GS, Henderson JT, Woo M, Soengas MS, et al. Differential requirement for caspase 9 in apoptotic pathways in vivo. Cell. 1998;94:339–52.
Marsden VS, Ekert PG, Van Delft M, Vaux DL, Adams JM, Strasser A. Bcl-2-regulated apoptosis and cytochrome c release can occur independently of both caspase-2 and caspase-9. J Cell Biol. 2004;165:775–80.
Spellicy CJ, Norris J, Bend R, Bupp C, Mester P, Reynolds T, et al. Key apoptotic genes APAF1 and CASP9 implicated in recurrent folate-resistant neural tube defects. Eur J Hum Genet. 2018;26:420–7.
Zhou X, Zeng W, Li H, Chen H, Wei G, Yang X, et al. Rare mutations in apoptosis related genes APAF1, CASP9, and CASP3 contribute to human neural tube defects. Cell Death Dis. 2018;9:43.
Li K, Li Y, Shelton JM, Richardson JA, Spencer E, Chen ZJ, et al. Cytochrome c deficiency causes embryonic lethality and attenuates stress-induced apoptosis. Cell. 2000;101:389–99.
Narisawa S, Hecht NB, Goldberg E, Boatright KM, Reed JC, Millán JL. Testis-specific cytochrome c-null mice produce functional sperm but undergo early testicular atrophy. Mol Cell Biol. 2002;22:5554–62.
Pinto M, Vempati UD, Diaz F, Peralta S, Moraes CT. Ablation of cytochrome c in Adult Forebrain Neurons Impairs Oxidative Phosphorylation Without Detectable Apoptosis. Mol Neurobiol. 2019;56:3722–35.
Hao Z, Duncan GS, Chang CC, Elia A, Fang M, Wakeham A, et al. Specific ablation of the apoptotic functions of cytochrome C reveals a differential requirement for cytochrome C and Apaf-1 in apoptosis. Cell. 2005;121:579–91.
Woo M, Hakem R, Soengas MS, Duncan GS, Shahinian A, Kägi D, et al. Essential contribution of caspase 3/CPP32 to apoptosis and its associated nuclear changes. Genes Dev. 1998;12:806–19.
Kuida K, Zheng TS, Na S, Kuan C, Yang D, Karasuyama H, et al. Decreased apoptosis in the brain and premature lethality in CPP32-deficient mice. Nature. 1996;384:368–72.
Leonard JR, Klocke BJ, D’Sa C, Flavell RA, Roth KA. Strain-dependent neurodevelopmental abnormalities in caspase-3-deficient mice. J Neuropathol Exp Neurol. 2002;61:673–7.
Roth KA, Kuan C, Haydar TF, D’Sa-Eipper C, Shindler KS, Zheng TS, et al. Epistatic and independent functions of caspase-3 and Bcl-X(L) in developmental programmed cell death. Proc Natl Acad Sci USA. 2000;97:466–71.
Matsumoto Y, Yamaguchi Y, Hamachi M, Nonomura K, Muramatsu Y, Yoshida H, et al. Apoptosis is involved in maintaining the character of the midbrain and the diencephalon roof plate after neural tube closure. Dev Biol. 2020;468:101–9.
Okamoto H, Shiraishi H, Yoshida H. Histological analyses of normally grown, fertile Apaf1-deficient mice. Cell Death Differ. 2006;13:668–71.
Lo SC, Scearce-Levie K, Sheng M. Characterization of social behaviors in caspase-3 deficient mice. Sci Rep. 2016;6:18335.
Lo SC, Wang Y, Weber M, Larson JL, Scearce-Levie K, Sheng M. Caspase-3 deficiency results in disrupted synaptic homeostasis and impaired attention control. J Neurosci. 2015;35:2118–32.
Takahashi K, Kamiya K, Urase K, Suga M, Takizawa T, Mori H, et al. Caspase-3-deficiency induces hyperplasia of supporting cells and degeneration of sensory cells resulting in the hearing loss. Brain Res. 2001;894:359–67.
Morishita H, Makishima T, Kaneko C, Lee YS, Segil N, Takahashi K, et al. Deafness due to degeneration of cochlear neurons in caspase-3-deficient mice. Biochem Biophys Res Commun. 2001;284:142–9.
Parker A, Hardisty-Hughes RE, Wisby L, Joyce S, Brown SD. Melody, an ENU mutation in Caspase 3, alters the catalytic cysteine residue and causes sensorineural hearing loss in mice. Mamm Genome. 2010;21:565–76.
Armstrong PA, Wood SJ, Shimizu N, Kuster K, Perachio A, Makishima T. Preserved otolith organ function in caspase-3-deficient mice with impaired horizontal semicircular canal function. Exp Brain Res. 2015;233:1825–35.
Makishima T, Hochman L, Armstrong P, Rosenberger E, Ridley R, Woo M, et al. Inner ear dysfunction in caspase-3 deficient mice. BMC Neurosci. 2011;12:102.
Suzuki T, Ichii O, Nakamura T, Horino T, Elewa YHA, Kon Y. Immune-associated renal disease found in caspase 3-deficient mice. Cell tissue Res. 2020;379:323–35.
Houde C, Banks KG, Coulombe N, Rasper D, Grimm E, Roy S, et al. Caspase-7 expanded function and intrinsic expression level underlies strain-specific brain phenotype of caspase-3-null mice. J Neurosci. 2004;24:9977–84.
Lakhani SA, Masud A, Kuida K, Porter GA Jr, Booth CJ, Mehal WZ, et al. Caspases 3 and 7: key mediators of mitochondrial events of apoptosis. Science. 2006;311:847–51.
McComb S, Chan PK, Guinot A, Hartmannsdottir H, Jenni S, Dobay MP, et al. Efficient apoptosis requires feedback amplification of upstream apoptotic signals by effector caspase-3 or -7. Sci Adv. 2019;5:eaau9433.
Walsh JG, Cullen SP, Sheridan C, Lüthi AU, Gerner C, Martin SJ. Executioner caspase-3 and caspase-7 are functionally distinct proteases. Proc Natl Acad Sci USA. 2008;105:12815–9.
Yoshida A, Kawata D, Shinotsuka N, Yoshida M, Yamaguchi Y, Miura M. Evidence for the involvement of caspases in establishing proper cerebrospinal fluid hydrodynamics. Neurosci Res. 2021;170:145–53.
Demon D, Van Damme P, Vanden Berghe T, Deceuninck A, Van Durme J, Verspurten J, et al. Proteome-wide substrate analysis indicates substrate exclusion as a mechanism to generate caspase-7 versus caspase-3 specificity. Mol Cell Proteom. 2009;8:2700–14.
Nozaki K, Maltez VI, Rayamajhi M, Tubbs AL, Mitchell JE, Lacey CA, et al. Caspase-7 activates ASM to repair gasdermin and perforin pores. Nature. 2022;606:960–7.
Okada H, Suh WK, Jin J, Woo M, Du C, Elia A, et al. Generation and characterization of Smac/DIABLO-deficient mice. Mol Cell Biol. 2002;22:3509–17.
Martins LM, Morrison A, Klupsch K, Fedele V, Moisoi N, Teismann P, et al. Neuroprotective role of the Reaper-related serine protease HtrA2/Omi revealed by targeted deletion in mice. Mol Cell Biol. 2004;24:9848–62.
Hui KK, Kanungo AK, Elia AJ, Henderson JT. Caspase-3 deficiency reveals a physiologic role for Smac/DIABLO in regulating programmed cell death. Cell Death Differ. 2011;18:1780–90.
Olayioye MA, Kaufmann H, Pakusch M, Vaux DL, Lindeman GJ, Visvader JE. XIAP-deficiency leads to delayed lobuloalveolar development in the mammary gland. Cell Death Differ. 2005;12:87–90.
Harlin H, Reffey SB, Duckett CS, Lindsten T, Thompson CB. Characterization of XIAP-deficient mice. Mol Cell Biol. 2001;21:3604–8.
Morrish E, Brumatti G, Silke J. Future therapeutic directions for smac-mimetics. Cells. 2020;9:406.
Prakash H, Albrecht M, Becker D, Kuhlmann T, Rudel T. Deficiency of XIAP leads to sensitization for Chlamydophila pneumoniae pulmonary infection and dysregulation of innate immune response in mice. J Biol Chem. 2010;285:20291–302.
Yabal M, Müller N, Adler H, Knies N, Groß CJ, Damgaard RB, et al. XIAP restricts TNF- and RIP3-dependent cell death and inflammasome activation. Cell Rep. 2014;7:1796–808.
Hsieh WC, Chuang YT, Chiang IH, Hsu SC, Miaw SC, Lai MZ. Inability to resolve specific infection generates innate immunodeficiency syndrome in Xiap−/− mice. Blood. 2014;124:2847–57.
Cardona M, López JA, Serafín A, Rongvaux A, Inserte J, García-Dorado D, et al. Executioner caspase-3 and 7 deficiency reduces myocyte number in the developing mouse heart. PLoS ONE. 2015;10:e0131411.
White MJ, Schoenwaelder SM, Josefsson EC, Jarman KE, Henley KJ, James C, et al. Caspase-9 mediates the apoptotic death of megakaryocytes and platelets, but is dispensable for their generation and function. Blood. 2012;119:4283–90.
van Delft MF, Smith DP, Lahoud MH, Huang DC, Adams JM. Apoptosis and non-inflammatory phagocytosis can be induced by mitochondrial damage without caspases. Cell Death Differ. 2010;17:821–32.
Woo M, Hakem R, Furlonger C, Hakem A, Duncan GS, Sasaki T, et al. Caspase-3 regulates cell cycle in B cells: a consequence of substrate specificity. Nat Immunol. 2003;4:1016–22.
Miura M, Chen XD, Allen MR, Bi Y, Gronthos S, Seo BM, et al. A crucial role of caspase-3 in osteogenic differentiation of bone marrow stromal stem cells. J Clin Investig. 2004;114:1704–13.
Tong H, Miyake Y, Mi-Ichi F, Iwakura Y, Hara H, Yoshida H. Apaf1 plays a negative regulatory role in T cell responses by suppressing activation of antigen-stimulated T cells. PLoS ONE. 2018;13:e0195119.
Hara H, Takeda A, Takeuchi M, Wakeham AC, Itié A, Sasaki M, et al. The apoptotic protease-activating factor 1-mediated pathway of apoptosis is dispensable for negative selection of thymocytes. J Immunol. 2002;168:2288–95.
Nagasaka A, Kawane K, Yoshida H, Nagata S. Apaf-1-independent programmed cell death in mouse development. Cell Death Differ. 2010;17:931–41.
Doerfler P, Forbush KA, Perlmutter RM. Caspase enzyme activity is not essential for apoptosis during thymocyte development. J Immunol. 2000;164:4071–9.
Izquierdo M, Grandien A, Criado LM, Robles S, Leonardo E, Albar JP, et al. Blocked negative selection of developing T cells in mice expressing the baculovirus p35 caspase inhibitor. EMBO J. 1999;18:156–66.
Lu EP, McLellan M, Ding L, Fulton R, Mardis ER, Wilson RK, et al. Caspase-9 is required for normal hematopoietic development and protection from alkylator-induced DNA damage in mice. Blood. 2014;124:3887–95.
Oppenheim RW, Blomgren K, Ethell DW, Koike M, Komatsu M, Prevette D, et al. Developing postmitotic mammalian neurons in vivo lacking Apaf-1 undergo programmed cell death by a caspase-independent, nonapoptotic pathway involving autophagy. J Neurosci. 2008;28:1490–7.
Oppenheim RW, Flavell RA, Vinsant S, Prevette D, Kuan CY, Rakic P. Programmed cell death of developing mammalian neurons after genetic deletion of caspases. J Neurosci. 2001;21:4752–60.
Yaginuma H, Shiraiwa N, Shimada T, Nishiyama K, Hong J, Wang S, et al. Caspase activity is involved in, but is dispensable for, early motoneuron death in the chick embryo cervical spinal cord. Mol Cell Neurosci. 2001;18:168–82.
Honarpour N, Tabuchi K, Stark JM, Hammer RE, Südhof TC, Parada LF, et al. Embryonic neuronal death due to neurotrophin and neurotransmitter deprivation occurs independent of Apaf-1. Neuroscience. 2001;106:263–74.
Aggarwal BB, Gupta SC, Kim JH. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood. 2012;119:651–65.
Strasser A, Jost PJ, Nagata S. The many roles of FAS receptor signaling in the immune system. Immunity. 2009;30:180–92.
Wajant H. The Fas signaling pathway: more than a paradigm. Science. 2002;296:1635–6.
Wallach D. The tumor necrosis factor family: family conventions and private idiosyncrasies. Cold Spring Harbor Perspect Biol. 2018;10:a028431.
von Karstedt S, Montinaro A, Walczak H. Exploring the TRAILs less travelled: TRAIL in cancer biology and therapy. Nat Rev Cancer. 2017;17:352–66.
Kischkel FC, Hellbardt S, Behrmann I, Germer M, Pawlita M, Krammer PH, et al. Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins form a death-inducing signaling complex (DISC) with the receptor. EMBO J. 1995;14:5579–88.
Boldin MP, Goncharov TM, Goltsev YV, Wallach D. Involvement of MACH, a novel MORT1/FADD-interacting protease, in Fas/APO-1- and TNF receptor-induced cell death. Cell. 1996;85:803–15.
Dickens LS, Powley IR, Hughes MA, MacFarlane M. The ‘complexities’ of life and death: death receptor signalling platforms. Exp cell Res. 2012;318:1269–77.
Muzio M, Chinnaiyan AM, Kischkel FC, O’Rourke K, Shevchenko A, Ni J, et al. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death-inducing signaling complex. Cell. 1996;85:817–27.
Boldin MP, Varfolomeev EE, Pancer Z, Mett IL, Camonis JH, Wallach D. A novel protein that interacts with the death domain of Fas/APO1 contains a sequence motif related to the death domain. J Biol Chem. 1995;270:7795–8.
Chinnaiyan AM, O’Rourke K, Tewari M, Dixit VM. FADD, a novel death domain-containing protein, interacts with the death domain of Fas and initiates apoptosis. Cell. 1995;81:505–12.
Kischkel FC, Lawrence DA, Chuntharapai A, Schow P, Kim KJ, Ashkenazi A. Apo2L/TRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity. 2000;12:611–20.
Scott FL, Stec B, Pop C, Dobaczewska MK, Lee JJ, Monosov E, et al. The Fas-FADD death domain complex structure unravels signalling by receptor clustering. Nature. 2009;457:1019–22.
Chan FK, Chun HJ, Zheng L, Siegel RM, Bui KL, Lenardo MJ. A domain in TNF receptors that mediates ligand-independent receptor assembly and signaling. Science. 2000;288:2351–4.
Fu Q, Fu TM, Cruz AC, Sengupta P, Thomas SK, Wang S, et al. StructuraL Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 death receptor. Mol Cell. 2016;61:602–13.
Brenner D, Blaser H, Mak TW. Regulation of tumour necrosis factor signalling: live or let die. Nat Rev Immunol. 2015;15:362–74.
Galluzzi L, Kepp O, Chan FK, Kroemer G. Necroptosis: Mechanisms and Relevance to Disease. Annu Rev Pathol. 2017;12:103–30.
Tummers B, Green DR. Caspase-8: regulating life and death. Immunol Rev. 2017;277:76–89.
Barnhart BC, Alappat EC, Peter ME. The CD95 type I/type II model. Semin Immunol. 2003;15:185–93.
Strasser A, Harris AW, Huang DC, Krammer PH, Cory S. Bcl-2 and Fas/APO-1 regulate distinct pathways to lymphocyte apoptosis. EMBO J. 1995;14:6136–47.
Li H, Zhu H, Xu CJ, Yuan J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell. 1998;94:491–501.
Luo X, Budihardjo I, Zou H, Slaughter C, Wang X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell. 1998;94:481–90.
Gross A, Yin XM, Wang K, Wei MC, Jockel J, Milliman C, et al. Caspase cleaved BID targets mitochondria and is required for cytochrome c release, while BCL-XL prevents this release but not tumor necrosis factor-R1/Fas death. J Biol Chem. 1999;274:1156–63.
Huang K, Zhang J, O’Neill KL, Gurumurthy CB, Quadros RM, Tu Y, et al. Cleavage by caspase 8 and mitochondrial membrane association activate the BH3-only protein bid during TRAIL-induced apoptosis. J Biol Chem. 2016;291:11843–51.
Lalaoui N, Boyden SE, Oda H, Wood GM, Stone DL, Chau D, et al. Mutations that prevent caspase cleavage of RIPK1 cause autoinflammatory disease. Nature. 2020;577:103–8.
Hayden MS, Ghosh S. Regulation of NF-κB by TNF family cytokines. Semin Immunol. 2014;26:253–66.
Bertheloot D, Latz E, Franklin BS. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cell Mol Immunol. 2021;18:1106–21.
Henry CM, Martin SJ. Caspase-8 acts in a non-enzymatic role as a scaffold for assembly of a pro-inflammatory “FADDosome” Complex upon TRAIL Stimulation. Mol Cell. 2017;65:715–729.e715.
Gibert B, Mehlen P. Dependence receptors and cancer: addiction to trophic ligands. Cancer Res. 2015;75:5171–5.
Mehlen P, Bredesen DE. Dependence receptors: from basic research to drug development. Sci Signal. 2011;4:mr2.
Brisset M, Grandin M, Bernet A, Mehlen P, Hollande F. Dependence receptors: new targets for cancer therapy. EMBO Mol Med. 2021;13:e14495.
Dillon CP, Oberst A, Weinlich R, Janke LJ, Kang TB, Ben-Moshe T, et al. Survival function of the FADD-CASPASE-8-cFLIP(L) complex. Cell Rep. 2012;1:401–7.
Dillon CP, Weinlich R, Rodriguez DA, Cripps JG, Quarato G, Gurung P, et al. RIPK1 blocks early postnatal lethality mediated by caspase-8 and RIPK3. Cell. 2014;157:1189–202.
Kaiser WJ, Daley-Bauer LP, Thapa RJ, Mandal P, Berger SB, Huang C, et al. RIP1 suppresses innate immune necrotic as well as apoptotic cell death during mammalian parturition. Proc Natl Acad Sci USA. 2014;111:7753–8.
Rickard JA, O’Donnell JA, Evans JM, Lalaoui N, Poh AR, Rogers T, et al. RIPK1 regulates RIPK3-MLKL-driven systemic inflammation and emergency hematopoiesis. Cell. 2014;157:1175–88.
Lynch DH, Watson ML, Alderson MR, Baum PR, Miller RE, Tough T, et al. The mouse Fas-ligand gene is mutated in gld mice and is part of a TNF family gene cluster. Immunity. 1994;1:131–6.
Roths JB, Murphy ED, Eicher EM. A new mutation, gld, that produces lymphoproliferation and autoimmunity in C3H/HeJ mice. J Exp Med. 1984;159:1–20.
Matsuzawa A, Moriyama T, Kaneko T, Tanaka M, Kimura M, Ikeda H, et al. A new allele of the lpr locus, lprcg, that complements the gld gene in induction of lymphadenopathy in the mouse. J Exp Med. 1990;171:519–31.
Adachi M, Suematsu S, Kondo T, Ogasawara J, Tanaka T, Yoshida N, et al. Targeted mutation in the Fas gene causes hyperplasia in peripheral lymphoid organs and liver. Nat Genet. 1995;11:294–300.
Anstee NS, Vandenberg CJ, Campbell KJ, Hughes PD, O’Reilly LA, Cory S. Overexpression of Mcl-1 exacerbates lymphocyte accumulation and autoimmune kidney disease in lpr mice. Cell Death Differ. 2017;24:397–408.
Hughes PD, Belz GT, Fortner KA, Budd RC, Strasser A, Bouillet P. Apoptosis regulators Fas and Bim cooperate in shutdown of chronic immune responses and prevention of autoimmunity. Immunity. 2008;28:197–205.
Karray S, Kress C, Cuvellier S, Hue-Beauvais C, Damotte D, Babinet C, et al. Complete loss of Fas ligand gene causes massive lymphoproliferation and early death, indicating a residual activity of gld allele. J Immunol. 2004;172:2118–25.
Davies MH, Eubanks JP, Powers MR. Increased retinal neovascularization in Fas ligand-deficient mice. Investig Ophthalmol Vis Sci. 2003;44:3202–10.
Schumann DM, Maedler K, Franklin I, Konrad D, Størling J, Böni-Schnetzler M, et al. The Fas pathway is involved in pancreatic beta cell secretory function. Proc Natl Acad Sci USA. 2007;104:2861–6.
Trumpi K, Steller EJ, de Leng WW, Raats DA, Nijman IJ, Morsink FH, et al. Mice lacking functional CD95-ligand display reduced proliferation of the intestinal epithelium without gross homeostatic alterations. Med Mol Morphol. 2016;49:110–8.
Fukuyama H, Adachi M, Suematsu S, Miwa K, Suda T, Yoshida N, et al. Transgenic expression of Fas in T cells blocks lymphoproliferation but not autoimmune disease in MRL-lpr mice. J Immunol. 1998;160:3805–11.
Komano H, Ikegami Y, Yokoyama M, Suzuki R, Yonehara S, Yamasaki Y, et al. Severe impairment of B cell function in lpr/lpr mice expressing transgenic Fas selectively on B cells. Int Immunol. 1999;11:1035–42.
Stranges PB, Watson J, Cooper CJ, Choisy-Rossi CM, Stonebraker AC, Beighton RA, et al. Elimination of antigen-presenting cells and autoreactive T cells by Fas contributes to prevention of autoimmunity. Immunity. 2007;26:629–41.
Rathmell JC, Cooke MP, Ho WY, Grein J, Townsend SE, Davis MM, et al. CD95 (Fas)-dependent elimination of self-reactive B cells upon interaction with CD4+ T cells. Nature. 1995;376:181–4.
Zhang JQ, Okumura C, McCarty T, Shin MS, Mukhopadhyay P, Hori M, et al. Evidence for selective transformation of autoreactive immature plasma cells in mice deficient in Fasl. J Exp Med. 2004;200:1467–78.
Peng SL, Robert ME, Hayday AC, Craft J. A tumor-suppressor function for Fas (CD95) revealed in T cell-deficient mice. J Exp Med. 1996;184:1149–54.
Sedger LM, Glaccum MB, Schuh JC, Kanaly ST, Williamson E, Kayagaki N, et al. Characterization of the in vivo function of TNF-alpha-related apoptosis-inducing ligand, TRAIL/Apo2L, using TRAIL/Apo2L gene-deficient mice. Eur J Immunol. 2002;32:2246–54.
Diehl GE, Yue HH, Hsieh K, Kuang AA, Ho M, Morici LA, et al. TRAIL-R as a negative regulator of innate immune cell responses. Immunity. 2004;21:877–89.
Finnberg N, Gruber JJ, Fei P, Rudolph D, Bric A, Kim SH, et al. DR5 knockout mice are compromised in radiation-induced apoptosis. Mol Cell Biol. 2005;25:2000–13.
Lehnert C, Weiswange M, Jeremias I, Bayer C, Grunert M, Debatin KM, et al. TRAIL-receptor costimulation inhibits proximal TCR signaling and suppresses human T cell activation and proliferation. J Immunol. 2014;193:4021–31.
McGrath EE, Marriott HM, Lawrie A, Francis SE, Sabroe I, Renshaw SA, et al. TNF-related apoptosis-inducing ligand (TRAIL) regulates inflammatory neutrophil apoptosis and enhances resolution of inflammation. J Leukoc Biol. 2011;90:855–65.
Sacks JA, Bevan MJ. TRAIL deficiency does not rescue impaired CD8+ T cell memory generated in the absence of CD4+ T cell help. J Immunol. 2008;180:4570–6.
Cretney E, Uldrich AP, Berzins SP, Strasser A, Godfrey DI, Smyth MJ. Normal thymocyte negative selection in TRAIL-deficient mice. J Exp Med. 2003;198:491–6.
Marino MW, Dunn A, Grail D, Inglese M, Noguchi Y, Richards E, et al. Characterization of tumor necrosis factor-deficient mice. Proc Natl Acad Sci USA. 1997;94:8093–8.
Pasparakis M, Alexopoulou L, Episkopou V, Kollias G. Immune and inflammatory responses in TNF alpha-deficient mice: a critical requirement for TNF alpha in the formation of primary B cell follicles, follicular dendritic cell networks and germinal centers, and in the maturation of the humoral immune response. J Exp Med. 1996;184:1397–411.
Pasparakis M, Alexopoulou L, Grell M, Pfizenmaier K, Bluethmann H, Kollias G. Peyer’s patch organogenesis is intact yet formation of B lymphocyte follicles is defective in peripheral lymphoid organs of mice deficient for tumor necrosis factor and its 55-kDa receptor. Proc Natl Acad Sci USA. 1997;94:6319–23.
Körner H, Cook M, Riminton DS, Lemckert FA, Hoek RM, Ledermann B, et al. Distinct roles for lymphotoxin-alpha and tumor necrosis factor in organogenesis and spatial organization of lymphoid tissue. Eur J Immunol. 1997;27:2600–9.
Oishi N, Chen J, Zheng HW, Hill K, Schacht J, Sha SH. Tumor necrosis factor-alpha-mutant mice exhibit high frequency hearing loss. J Assoc Res Otolaryngology. 2013;14:801–11.
Pfeffer K, Matsuyama T, Kündig TM, Wakeham A, Kishihara K, Shahinian A, et al. Mice deficient for the 55 kd tumor necrosis factor receptor are resistant to endotoxic shock, yet succumb to L. monocytogenes infection. Cell. 1993;73:457–67.
Rothe J, Lesslauer W, Lötscher H, Lang Y, Koebel P, Köntgen F, et al. Mice lacking the tumour necrosis factor receptor 1 are resistant to TNF-mediated toxicity but highly susceptible to infection by Listeria monocytogenes. Nature. 1993;364:798–802.
Kubota T, McTiernan CF, Frye CS, Demetris AJ, Feldman AM. Cardiac-specific overexpression of tumor necrosis factor-alpha causes lethal myocarditis in transgenic mice. J Card Fail. 1997;3:117–24.
Kubota T, McTiernan CF, Frye CS, Slawson SE, Lemster BH, Koretsky AP, et al. Dilated cardiomyopathy in transgenic mice with cardiac-specific overexpression of tumor necrosis factor-alpha. Circ Res. 1997;81:627–35.
Lacey D, Hickey P, Arhatari BD, O’Reilly LA, Rohrbeck L, Kiriazis H, et al. Spontaneous retrotransposon insertion into TNF 3’UTR causes heart valve disease and chronic polyarthritis. Proc Natl Acad Sci USA. 2015;112:9698–703.
Sedger LM, Katewa A, Pettersen AK, Osvath SR, Farrell GC, Stewart GJ, et al. Extreme lymphoproliferative disease and fatal autoimmune thrombocytopenia in FasL and TRAIL double-deficient mice. Blood. 2010;115:3258–68.
Körner H, Cretney E, Wilhelm P, Kelly JM, Röllinghoff M, Sedgwick JD, et al. Tumor necrosis factor sustains the generalized lymphoproliferative disorder (gld) phenotype. J Exp Med. 2000;191:89–96.
Meynier S, Rieux-Laucat F. FAS and RAS related apoptosis defects: from autoimmunity to leukemia. Immunol Rev. 2019;287:50–61.
Rieux-Laucat F, Magérus-Chatinet A, Neven B. The autoimmune lymphoproliferative syndrome with defective FAS or FAS-ligand functions. J Clin Immunol. 2018;38:558–68.
Del-Rey M, Ruiz-Contreras J, Bosque A, Calleja S, Gomez-Rial J, Roldan E, et al. A homozygous Fas ligand gene mutation in a patient causes a new type of autoimmune lymphoproliferative syndrome. Blood. 2006;108:1306–12.
Fisher GH, Rosenberg FJ, Straus SE, Dale JK, Middleton LA, Lin AY, et al. Dominant interfering Fas gene mutations impair apoptosis in a human autoimmune lymphoproliferative syndrome. Cell. 1995;81:935–46.
Magerus-Chatinet A, Stolzenberg MC, Loffredo MS, Neven B, Schaffner C, Ducrot N, et al. FAS-L, IL-10, and double-negative CD4- CD8- TCR alpha/beta+ T cells are reliable markers of autoimmune lymphoproliferative syndrome (ALPS) associated with FAS loss of function. Blood. 2009;113:3027–30.
Rensing-Ehl A, Völkl S, Speckmann C, Lorenz MR, Ritter J, Janda A, et al. Abnormally differentiated CD4+ or CD8+ T cells with phenotypic and genetic features of double negative T cells in human Fas deficiency. Blood. 2014;124:851–60.
Price S, Shaw PA, Seitz A, Joshi G, Davis J, Niemela JE, et al. Natural history of autoimmune lymphoproliferative syndrome associated with FAS gene mutations. Blood. 2014;123:1989–99.
Bi LL, Pan G, Atkinson TP, Zheng L, Dale JK, Makris C, et al. Dominant inhibition of Fas ligand-mediated apoptosis due to a heterozygous mutation associated with autoimmune lymphoproliferative syndrome (ALPS) Type Ib. BMC Med Genet. 2007;8:41.
Venkataraman G, McClain KL, Pittaluga S, Rao VK, Jaffe ES. Development of disseminated histiocytic sarcoma in a patient with autoimmune lymphoproliferative syndrome and associated Rosai-Dorfman disease. Am J Surg Pathol. 2010;34:589–94.
Haas SL, Lohse P, Schmitt WH, Hildenbrand R, Karaorman M, Singer MV, et al. Severe TNF receptor-associated periodic syndrome due to 2 TNFRSF1A mutations including a new F60V substitution. Gastroenterology. 2006;130:172–8.
Tsuji S, Matsuzaki H, Iseki M, Nagasu A, Hirano H, Ishihara K, et al. Functional analysis of a novel G87V TNFRSF1A mutation in patients with TNF receptor-associated periodic syndrome. Clin Exp Immunol. 2019;198:416–29.
McDermott MF, Aksentijevich I, Galon J, McDermott EM, Ogunkolade BW, Centola M, et al. Germline mutations in the extracellular domains of the 55 kDa TNF receptor, TNFR1, define a family of dominantly inherited autoinflammatory syndromes. Cell. 1999;97:133–44.
Yeh WC, de la Pompa JL, McCurrach ME, Shu HB, Elia AJ, Shahinian A, et al. FADD: essential for embryo development and signaling from some, but not all, inducers of apoptosis. Science. 1998;279:1954–8.
Imtiyaz HZ, Zhou X, Zhang H, Chen D, Hu T, Zhang J. The death domain of FADD is essential for embryogenesis, lymphocyte development, and proliferation. J Biol Chem. 2009;284:9917–26.
Zhang H, Zhou X, McQuade T, Li J, Chan FK, Zhang J. Functional complementation between FADD and RIP1 in embryos and lymphocytes. Nature. 2011;471:373–6.
Varfolomeev EE, Schuchmann M, Luria V, Chiannilkulchai N, Beckmann JS, Mett IL, et al. Targeted disruption of the mouse Caspase 8 gene ablates cell death induction by the TNF receptors, Fas/Apo1, and DR3 and is lethal prenatally. Immunity. 1998;9:267–76.
Sakamaki K, Inoue T, Asano M, Sudo K, Kazama H, Sakagami J, et al. Ex vivo whole-embryo culture of caspase-8-deficient embryos normalize their aberrant phenotypes in the developing neural tube and heart. Cell Death Differ. 2002;9:1196–206.
Yeh WC, Itie A, Elia AJ, Ng M, Shu HB, Wakeham A, et al. Requirement for Casper (c-FLIP) in regulation of death receptor-induced apoptosis and embryonic development. Immunity. 2000;12:633–42.
Chen NJ, Chio II, Lin WJ, Duncan G, Chau H, Katz D, et al. Beyond tumor necrosis factor receptor: TRADD signaling in toll-like receptors. Proc Natl Acad Sci USA. 2008;105:12429–34.
Ermolaeva MA, Michallet MC, Papadopoulou N, Utermöhlen O, Kranidioti K, Kollias G, et al. Function of TRADD in tumor necrosis factor receptor 1 signaling and in TRIF-dependent inflammatory responses. Nat Immunol. 2008;9:1037–46.
Pobezinskaya YL, Kim YS, Choksi S, Morgan MJ, Li T, Liu C, et al. The function of TRADD in signaling through tumor necrosis factor receptor 1 and TRIF-dependent Toll-like receptors. Nat Immunol. 2008;9:1047–54.
Kelliher MA, Grimm S, Ishida Y, Kuo F, Stanger BZ, Leder P. The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity. 1998;8:297–303.
Roderick JE, Hermance N, Zelic M, Simmons MJ, Polykratis A, Pasparakis M, et al. Hematopoietic RIPK1 deficiency results in bone marrow failure caused by apoptosis and RIPK3-mediated necroptosis. Proc Natl Acad Sci USA. 2014;111:14436–41.
Moulin M, Anderton H, Voss AK, Thomas T, Wong WW, Bankovacki A, et al. IAPs limit activation of RIP kinases by TNF receptor 1 during development. EMBO J. 2012;31:1679–91.
Heard KN, Bertrand MJ, Barker PA. cIAP2 supports viability of mice lacking cIAP1 and XIAP. EMBO J. 2015;34:2393–5.
Zhao Q, Yu X, Zhang H, Liu Y, Zhang X, Wu X, et al. RIPK3 mediates necroptosis during embryonic development and postnatal inflammation in Fadd-deficient mice. Cell Rep. 2017;19:798–808.
Newton K, Wickliffe KE, Dugger DL, Maltzman A, Roose-Girma M, Dohse M, et al. Cleavage of RIPK1 by caspase-8 is crucial for limiting apoptosis and necroptosis. Nature. 2019;574:428–31.
Zhang X, Dowling JP, Zhang J. RIPK1 can mediate apoptosis in addition to necroptosis during embryonic development. Cell Death Dis. 2019;10:245.
Anderton H, Bandala-Sanchez E, Simpson DS, Rickard JA, Ng AP, Di Rago L, et al. RIPK1 prevents TRADD-driven, but TNFR1 independent, apoptosis during development. Cell Death Differ. 2019;26:877–89.
Dowling JP, Alsabbagh M, Del Casale C, Liu ZG, Zhang J. TRADD regulates perinatal development and adulthood survival in mice lacking RIPK1 and RIPK3. Nat Commun. 2019;10:705.
Kang TB, Oh GS, Scandella E, Bolinger B, Ludewig B, Kovalenko A, et al. Mutation of a self-processing site in caspase-8 compromises its apoptotic but not its nonapoptotic functions in bacterial artificial chromosome-transgenic mice. J Immunol. 2008;181:2522–32.
Fritsch M, Günther SD, Schwarzer R, Albert MC, Schorn F, Werthenbach JP, et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature. 2019;575:683–7.
Tummers B, Mari L, Guy CS, Heckmann BL, Rodriguez DA, Rühl S, et al. Caspase-8-dependent inflammatory responses are controlled by its adaptor, FADD, and necroptosis. Immunity. 2020;52:994–1006.e1008.
Newton K, Wickliffe KE, Maltzman A, Dugger DL, Reja R, Zhang Y, et al. Activity of caspase-8 determines plasticity between cell death pathways. Nature. 2019;575:679–82.
Fan C, Pu W, Wu X, Zhang X, He L, Zhou B, et al. Lack of FADD in Tie-2 expressing cells causes RIPK3-mediated embryonic lethality. Cell Death Dis. 2016;7:e2351.
Zörnig M, Hueber AO, Evan G. p53-dependent impairment of T-cell proliferation in FADD dominant-negative transgenic mice. Curr Biol. 1998;8:467–70.
Zhang J, Cado D, Chen A, Kabra NH, Winoto A. Fas-mediated apoptosis and activation-induced T-cell proliferation are defective in mice lacking FADD/Mort1. Nature. 1998;392:296–300.
Zhang X, Dong X, Wang H, Li J, Yang B, Zhang J, et al. FADD regulates thymocyte development at the β-selection checkpoint by modulating Notch signaling. Cell Death Dis. 2014;5:e1273.
Newton K, Harris AW, Strasser A. FADD/MORT1 regulates the pre-TCR checkpoint and can function as a tumour suppressor. EMBO J. 2000;19:931–41.
Kabra NH, Kang C, Hsing LC, Zhang J, Winoto A. T cell-specific FADD-deficient mice: FADD is required for early T cell development. Proc Natl Acad Sci USA. 2001;98:6307–12.
Zhang Y, Rosenberg S, Wang H, Imtiyaz HZ, Hou YJ, Zhang J. Conditional Fas-associated death domain protein (FADD): GFP knockout mice reveal FADD is dispensable in thymic development but essential in peripheral T cell homeostasis. J Immunol. 2005;175:3033–44.
Osborn SL, Diehl G, Han SJ, Xue L, Kurd N, Hsieh K, et al. Fas-associated death domain (FADD) is a negative regulator of T-cell receptor-mediated necroptosis. Proc Natl Acad Sci USA. 2010;107:13034–9.
Zhang XY, Yang BY, Wang JY, Mo X, Zhang J, Hua ZC. FADD is essential for glucose uptake and survival of thymocytes. Biochem Biophys Res Commun. 2014;451:202–7.
Walsh CM, Wen BG, Chinnaiyan AM, O’Rourke K, Dixit VM, Hedrick SM. A role for FADD in T cell activation and development. Immunity. 1998;8:439–49.
Newton K, Kurts C, Harris AW, Strasser A. Effects of a dominant interfering mutant of FADD on signal transduction in activated T cells. Curr Biol. 2001;11:273–6.
Imtiyaz HZ, Rosenberg S, Zhang Y, Rahman ZS, Hou YJ, Manser T, et al. The Fas-associated death domain protein is required in apoptosis and TLR-induced proliferative responses in B cells. J Immunol. 2006;176:6852–61.
Salmena L, Lemmers B, Hakem A, Matysiak-Zablocki E, Murakami K, Au PY, et al. Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity. Genes Dev. 2003;17:883–95.
Beisner DR, Ch’en IL, Kolla RV, Hoffmann A, Hedrick SM. Cutting edge: innate immunity conferred by B cells is regulated by caspase-8. J Immunol. 2005;175:3469–73.
Lemmers B, Salmena L, Bidère N, Su H, Matysiak-Zablocki E, Murakami K, et al. Essential role for caspase-8 in Toll-like receptors and NFkappaB signaling. J Biol Chem. 2007;282:7416–23.
Zhang N, He YW. An essential role for c-FLIP in the efficient development of mature T lymphocytes. J Exp Med. 2005;202:395–404.
Chau H, Wong V, Chen NJ, Huang HL, Lin WJ, Mirtsos C, et al. Cellular FLICE-inhibitory protein is required for T cell survival and cycling. J Exp Med. 2005;202:405–13.
Zhang H, Rosenberg S, Coffey FJ, He YW, Manser T, Hardy RR, et al. A role for cFLIP in B cell proliferation and stress MAPK regulation. J Immunol. 2009;182:207–15.
Kennedy NJ, Kataoka T, Tschopp J, Budd RC. Caspase activation is required for T cell proliferation. J Exp Med. 1999;190:1891–6.
Bohgaki T, Mozo J, Salmena L, Matysiak-Zablocki E, Bohgaki M, Sanchez O, et al. Caspase-8 inactivation in T cells increases necroptosis and suppresses autoimmunity in Bim−/− mice. J Cell Biol. 2011;195:277–91.
Salmena L, Hakem R. Caspase-8 deficiency in T cells leads to a lethal lymphoinfiltrative immune disorder. J Exp Med. 2005;202:727–32.
Rosenberg S, Zhang H, Zhang J. FADD deficiency impairs early hematopoiesis in the bone marrow. J Immunol. 2011;186:203–13.
Pellegrini M, Bath S, Marsden VS, Huang DC, Metcalf D, Harris AW, et al. FADD and caspase-8 are required for cytokine-induced proliferation of hemopoietic progenitor cells. Blood. 2005;106:1581–9.
Schock SN, Young JA, He TH, Sun Y, Winoto A. Deletion of FADD in macrophages and granulocytes results in RIP3- and MyD88-dependent systemic inflammation. PLoS ONE. 2015;10:e0124391.
Cuda CM, Misharin AV, Khare S, Saber R, Tsai F, Archer AM, et al. Conditional deletion of caspase-8 in macrophages alters macrophage activation in a RIPK-dependent manner. Arthritis Res Ther. 2015;17:291.
Vitale I, Manic G, Coussens LM, Kroemer G, Galluzzi L. Macrophages and metabolism in the tumor microenvironment. Cell Metab. 2019;30:36–50.
Cuda CM, Misharin AV, Gierut AK, Saber R, Haines GK 3rd, Hutcheson J, et al. Caspase-8 acts as a molecular rheostat to limit RIPK1- and MyD88-mediated dendritic cell activation. J Immunol. 2014;192:5548–60.
Huang QQ, Perlman H, Birkett R, Doyle R, Fang D, Haines GK, et al. CD11c-mediated deletion of Flip promotes autoreactivity and inflammatory arthritis. Nat Commun. 2015;6:7086.
Wu YJ, Wu YH, Mo ST, Hsiao HW, He YW, Lai MZ. Cellular FLIP inhibits myeloid cell activation by suppressing selective innate signaling. J Immunol. 2015;195:2612–23.
Ch’en IL, Tsau JS, Molkentin JD, Komatsu M, Hedrick SM. Mechanisms of necroptosis in T cells. J Exp Med. 2011;208:633–41.
Bell BD, Leverrier S, Weist BM, Newton RH, Arechiga AF, Luhrs KA, et al. FADD and caspase-8 control the outcome of autophagic signaling in proliferating T cells. Proc Natl Acad Sci USA. 2008;105:16677–82.
Ch’en IL, Beisner DR, Degterev A, Lynch C, Yuan J, Hoffmann A, et al. Antigen-mediated T cell expansion regulated by parallel pathways of death. Proc Natl Acad Sci USA. 2008;105:17463–8.
Bolze A, Byun M, McDonald D, Morgan NV, Abhyankar A, Premkumar L, et al. Whole-exome-sequencing-based discovery of human FADD deficiency. Am J Hum Genet. 2010;87:873–81.
Kuehn HS, Caminha I, Niemela JE, Rao VK, Davis J, Fleisher TA, et al. FAS haploinsufficiency is a common disease mechanism in the human autoimmune lymphoproliferative syndrome. J Immunol. 2011;186:6035–43.
Kohn LA, Long JD, Trope EC, Kuo CY. Novel compound heterozygote variations in FADD identified to cause FAS-associated protein with death domain deficiency. J Clin Immunol. 2020;40:658–61.
Savic S, Parry D, Carter C, Johnson C, Logan C, Gutierrez BM, et al. A new case of Fas-associated death domain protein deficiency and update on treatment outcomes. J allergy Clin Immunol. 2015;136:502–505.e504.
Chun HJ, Zheng L, Ahmad M, Wang J, Speirs CK, Siegel RM, et al. Pleiotropic defects in lymphocyte activation caused by caspase-8 mutations lead to human immunodeficiency. Nature. 2002;419:395–9.
Wang J, Zheng L, Lobito A, Chan FK, Dale J, Sneller M, et al. Inherited human Caspase 10 mutations underlie defective lymphocyte and dendritic cell apoptosis in autoimmune lymphoproliferative syndrome type II. Cell. 1999;98:47–58.
Martínez-Feito A, Melero J, Mora-Díaz S, Rodríguez-Vigil C, Elduayen R, González-Granado LI, et al. Autoimmune lymphoproliferative syndrome due to somatic FAS mutation (ALPS-sFAS) combined with a germline caspase-10 (CASP10) variation. Immunobiology. 2016;221:40–47.
Dechant MJ, Scheuerpflug CG, Pauly E, van der Werff Ten Bosch J, Debatin KM, Fellenberg J. Screening, identification, and functional analysis of three novel missense mutations in the TRADD gene in children with ALL and ALPS. Pediatr Blood Cancer. 2008;51:616–20.
Lehle AS, Farin HF, Marquardt B, Michels BE, Magg T, Li Y, et al. Intestinal inflammation and dysregulated immunity in patients with inherited caspase-8 deficiency. Gastroenterology. 2019;156:275–8.
Niemela J, Kuehn HS, Kelly C, Zhang M, Davies J, Melendez J, et al. Caspase-8 deficiency presenting as late-onset multi-organ lymphocytic infiltration with granulomas in two adult siblings. J Clin Immunol. 2015;35:348–55.
Kanderova V, Grombirikova H, Zentsova I, Reblova K, Klocperk A, Fejtkova M, et al. Lymphoproliferation, immunodeficiency and early-onset inflammatory bowel disease associated with a novel mutation in Caspase 8. Haematologica. 2019;104:e32–e34.
Piao X, Komazawa-Sakon S, Nishina T, Koike M, Piao JH, Ehlken H, et al. c-FLIP maintains tissue homeostasis by preventing apoptosis and programmed necrosis. Sci Signal. 2012;5:ra93.
Panayotova-Dimitrova D, Feoktistova M, Ploesser M, Kellert B, Hupe M, Horn S, et al. cFLIP regulates skin homeostasis and protects against TNF-induced keratinocyte apoptosis. Cell Rep. 2013;5:397–408.
Feoktistova M, Makarov R, Leverkus M, Yazdi AS, Panayotova-Dimitrova D. TNF is partially required for cell-death-triggered skin inflammation upon acute loss of cFLIP. Int J Mol Sci. 2020;21:8859.
Wittkopf N, Günther C, Martini E, He G, Amann K, He YW, et al. Cellular FLICE-like inhibitory protein secures intestinal epithelial cell survival and immune homeostasis by regulating caspase-8. Gastroenterology. 2013;145:1369–79.
Gehrke N, Garcia-Bardon D, Mann A, Schad A, Alt Y, Wörns MA, et al. Acute organ failure following the loss of anti-apoptotic cellular FLICE-inhibitory protein involves activation of innate immune receptors. Cell Death Differ. 2015;22:826–37.
Bonnet MC, Preukschat D, Welz PS, van Loo G, Ermolaeva MA, Bloch W, et al. The adaptor protein FADD protects epidermal keratinocytes from necroptosis in vivo and prevents skin inflammation. Immunity. 2011;35:572–82.
Welz PS, Wullaert A, Vlantis K, Kondylis V, Fernández-Majada V, Ermolaeva M, et al. FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation. Nature. 2011;477:330–4.
Kovalenko A, Kim JC, Kang TB, Rajput A, Bogdanov K, Dittrich-Breiholz O, et al. Caspase-8 deficiency in epidermal keratinocytes triggers an inflammatory skin disease. J Exp Med. 2009;206:2161–77.
Günther C, Martini E, Wittkopf N, Amann K, Weigmann B, Neumann H, et al. Caspase-8 regulates TNF-α-induced epithelial necroptosis and terminal ileitis. Nature. 2011;477:335–9.
Li C, Lasse S, Lee P, Nakasaki M, Chen SW, Yamasaki K, et al. Development of atopic dermatitis-like skin disease from the chronic loss of epidermal caspase-8. Proc Natl Acad Sci USA. 2010;107:22249–54.
Kaden-Volynets V, Günther C, Zimmermann J, Beisner J, Becker C, Bischoff SC. Deletion of the Casp8 gene in mice results in ileocolitis, gut barrier dysfunction, and malassimilation, which can be partially attenuated by inulin or sodium butyrate. Am J Physiol Gastrointest Liver Physiol. 2019;317:G493–g507.
Weinlich R, Oberst A, Dillon CP, Janke LJ, Milasta S, Lukens JR, et al. Protective roles for caspase-8 and cFLIP in adult homeostasis. Cell Rep. 2013;5:340–8.
Stolzer I, Kaden-Volynets V, Ruder B, Letizia M, Bittel M, Rausch P, et al. Environmental microbial factors determine the pattern of inflammatory lesions in a murine model of Crohn’s disease-like inflammation. Inflamm Bowel Dis. 2020;26:66–79.
Kaemmerer E, Kuhn P, Schneider U, Jeon MK, Klaus C, Schiffer M, et al. Intestinal genetic inactivation of caspase-8 diminishes migration of enterocytes. World J Gastroenterol. 2015;21:4499–508.
Tisch N, Mogler C, Stojanovic A, Luck R, Korhonen EA, Ellerkmann A, et al. Caspase-8 in endothelial cells maintains gut homeostasis and prevents small bowel inflammation in mice. EMBO Mol Med. 2022;14:e14121.
Schwarzer R, Jiao H, Wachsmuth L, Tresch A, Pasparakis M. FADD and caspase-8 regulate gut homeostasis and inflammation by controlling MLKL- and GSDMD-mediated death of intestinal epithelial cells. Immunity. 2020;52:978–993.e976.
Karki R, Kanneganti TD. Diverging inflammasome signals in tumorigenesis and potential targeting. Nat Rev Cancer. 2019;19:197–214.
Funding
I. Vitale is and has been supported by the Associazione Italiana per la Ricerca sul Cancro (AIRC, IG 2017 #20417 and IG 2022 #27685) and by a startup grant from the Italian Institute for Genomic Medicine (Candiolo, Turin, Italy) and Compagnia di San Paolo (Torino, Italy). M. Piacentini, G. Melino, S. Melino, G. Ciliberto are supported by the Ministro dell’Università (Italy) progetto Heal Italia PE6. L. Galluzzi is/has been supported (as a PI unless otherwise indicated) by two Breakthrough Level 2 grants from the US DoD BCRP (#BC180476P1; #BC210945), by a Transformative Breast Cancer Consortium Grant from the US DoD BCRP (#W81XWH2120034, PI: Formenti), by a U54 grant from NIH/NCI (#CA274291, PI: Deasy, Formenti, Weichselbaum), by the 2019 Laura Ziskin Prize in Translational Research (#ZP-6177, PI: Formenti) from the Stand Up to Cancer (SU2C), by a Mantle Cell Lymphoma Research Initiative (MCL-RI, PI: Chen-Kiang) grant from the Leukemia and Lymphoma Society (LLS), by a Rapid Response Grant from the Functional Genomics Initiative (New York, US), by startup funds from the Dept. of Radiation Oncology at Weill Cornell Medicine (New York, US), by industrial collaborations with Lytix Biopharma (Oslo, Norway), Promontory (New York, US) and Onxeo (Paris, France), as well as by donations from Promontory (New York, US), the Luke Heller TECPR2 Foundation (Boston, US), Sotio a.s. (Prague, Czech Republic), Lytix Biopharma (Oslo, Norway), Onxeo (Paris, France), Ricerchiamo (Brescia, Italy), and Noxopharm (Chatswood, Australia). G. Kroemer is supported by the Ligue contre le Cancer (équipe labellisée); Agence National de la Recherche (ANR) – Projets blancs; AMMICa US23/CNRS UMS3655; Association pour la recherche sur le cancer (ARC); Cancéropôle Ile-de-France; European Research Council Advanced Investigator Grand “ICD-Cancer”, Fondation pour la Recherche Médicale (FRM); a donation by Elior; Equipex Onco-Pheno-Screen; European Joint Programme on Rare Diseases (EJPRD); European Research Council (ICD-Cancer), European Union Horizon 2020 Projects Oncobiome and Crimson; Fondation Carrefour; Institut National du Cancer (INCa); Institut Universitaire de France; LabEx Immuno-Oncology (ANR-18-IDEX-0001); a Cancer Research ASPIRE Award from the Mark Foundation; the RHU Immunolife; Seerave Foundation; SIRIC Stratified Oncology Cell DNA Repair and Tumor Immune Elimination (SOCRATE); and SIRIC Cancer Research and Personalized Medicine (CARPEM)
Author information
Authors and Affiliations
Contributions
L. Galluzzi and I. Vitale conceived the review and wrote the first version of the manuscript with constructive input from all authors. E. Guilbaud, C. Galassi and G. Manic prepared display items under the supervision of L. Galluzzi and I. Vitale. I. Vitale, F. Pietrocola and L. Galluzzi addressed requests from the Reviewers and Editors of Cell Death and Differentiation. All authors approved the final version of the article and figures.
Corresponding authors
Ethics declarations
Competing interests
All Editorial Board Members of Cell Death Differentiation, Cell Death Disease, or Cell Death Discovery are included among the authors. A. Degterev is shareholder in Denali Therapeutics. A. D. Garg has received renumeration, honorarium or consultancy fees from Boehringer Ingelheim, Miltenyi Biotec or IsoPlexis. A. Strasser and P. E. Czabotar (PEC) are employees of the Walter and Eliza Hall Institute, which has an agreement with Genentech and AbbVie and receives milestone and royalty payments related to venetoclax. Employees of the Walter and Eliza Hall Institute may be eligible for financial benefits related to these payments. PEC receives such a financial benefit as a result of previous research related to venetoclax. B. De Strooper (BDS) has no COI with the content of this paper. Possible perceived COI: BDS is or has been a consultant for Eli Lilly, Biogen, Janssen Pharmaceutica, Eisai, AbbVie and other companies. BDS is also a scientific founder of Augustine Therapeutics and a scientific founder and stockholder of Muna therapeutics. B. R. Stockwell is an inventor on patents and patent applications involving small molecule drug discovery, ferroptosis, and the antibody 3F3-FMA; co-founded and serves as a consultant to Exarta Therapeutics, and ProJenX Inc.; holds equity in Sonata Therapeutics, and serves as a consultant to Weatherwax Biotechnologies Corporation and Akin Gump Strauss Hauer & Feld LLP, and receives research funding from Sumitomo Dainippon Pharma Oncology. C. M. Troy has the following patent applications US20200164026, US20190142915, US20150165061, US20140024597. US2020058683, WO2018013519, WO/2020/223212. D. C. Rubinsztein serves as a consultant for Alladdin Healthcare Technologies Ltd., Mindrank AI, Nido Biosciences, Drishti Discoveries and PAQ Therapeutics. D. J. Murphy receives funding from the Merck Group (Darmstadt, Germany) & Puma Biotechnology (Los Angeles, CA, USA). D. Ofengeim is employed at SANOFI. JTO serves as a consultant for Anji Pharmaceuticals. J. T. Opferman receives research funding from AbbVie. D. W. Andrews receives funding for research in apoptosis from Amylyx. F Di Virgilio is Member of the Scientific Advisory Board (SAB) of Biosceptre Ltd (UK), and a consultant with Axxam SpA (Italy). F. Madeo has financial interest in TLL, The Longevity Labs and Samsara Therapeutics. P. Mehlen is founder and shareholder of NETRIS Pharma. G. A. Calin is the scientific founder of Ithax Pharmaceuticals. G. L. Kelly (GLK) is an employee of the Walter and Eliza Hall Institute which receives milestone and royalty payments related to venetoclax. GLK has received research funding from Servier. J. R. Cubillos-Ruiz is a scientific consultant for NextRNA Therapeutics, Inc. and Autoimmunity Biologic Solutions, Inc. K. Rajalingam is a founder and MD of KH Biotec GmbH. CVR is a scientific founder and member of the Scientific Advisory Board (SAB) of Surface Oncology, a member of Janssen Immunology SAB, and a consultant for the Roche Immunology Incubator. C. V. Rothlin has received grant support from Mirati Therapeutics. K. Schroder is a co-inventor on patent applications for NLRP3 inhibitors which have been licensed to Inflazome Ltd, a company headquartered in Dublin, Ireland. Inflazome is developing drugs that target the NLRP3 inflammasome to address unmet clinical needs in inflammatory disease. K. Schroder served on the Scientific Advisory Board of Inflazome in 2016–2017, and serves as a consultant to Quench Bio, USA and Novartis, Switzerland. L. Altucci receives funding for research from Merck and has shares in Epic SRL and CIRCE SRL. B. Joseph is cofounder of CERVO Therapeutics AB. M. Carbone received donations from the UH Foundation through donations from: the Riviera United-4-a Cure, the Melohn Family Endowment, the Honeywell International Inc., the Germaine Hope Brennan Foundation, and the Maurice and Joanna Sullivan Family Foundation. M. Carbone has a patent issued for BAP1. M. Carbone and two patents issued for HMGB1. M. Carbone is a board-certified pathologist who provides consultation for pleural pathology, including medical-legal. M. Conrad is a co-founder and shareholder of ROSCUE Therapeutics GmbH. M. Karin received support from Jansen Pharmaceuticals, Merck and Gossamer Bioscience. M. Karin is founder and member of SAB, Elgia Pharmaceuticals. M. Pagano is a scientific cofounder of SEED Therapeutics; receives research funding from and is a shareholder in Kymera Therapeutics; and is a consultant for, a member of the scientific advisory board of, and has financial interests in CullGen, SEED Therapeutics, Triana Biomedicines, and Umbra Therapeutics. M. G. Vander Heiden is a scientific advisor for Agios Pharmaceuticals, iTeos Therapeutics, Sage Therapeutics, Droia Ventures, and Auron Therapeutics. M. Oren is a consultant for Quintrigen. O. Kepp is a co-founder of Samsara Therapeutics. P. J. Jost has had a consulting or advisory role, received honoraria, research funding, and/or travel/accommodation expenses from: Ariad, Abbvie, Bayer, Boehringer, Novartis, Pfizer, Servier, Roche, BMS and Celgene, Pierre Fabre, Janssen/Johnson&Johnson, MSD. R. B. Damgaard is a scientific advisor for Immagene B.V., Amsterdam, Netherlands. R. I. Aqeilan serves as consultant for Mahzi Therapeutics. R. J. DeBerardinis is a founder and advisor at Atavistik Bio and serves on the Scientific Advisory Boards of Agios Pharmaceuticals, Vida Ventures, Droia Ventures and Nirogy Therapeutics. R. N. Ktsis is Co-founder of ASPIDA Therapeutics Inc. S. A. Lipton (SAL) discloses that he is the named inventor on worldwide patents for the use of memantine and derivatives for the treatment of neurodegenerative disorders. As per the rules of Harvard University, SAL participates in a Royalty Sharing Agreement with is former institutions, Boston Children’s Hospital/Harvard Medical School, which licensed these patents to Forest Laboratories and Allergan, now owned by Abbvie. SAL is also a founder of EuMentis Therapeutics, Inc., Adamas Pharmaceuticals, Inc. (now owned by Supernus Pharmaceuticals, Inc.), and a consultant to SNO bio, Inc., Engine Biosciences, Ventus Therapeutics, Inc., Eisai, inc., and Takeda Pharmaceuticals, Inc. S. Ghosh has received grant support from Mirati Therapeutics. S. J. Dixon is a co-founder of Prothegen Inc., and a scientific advisor to Ferro Therapeutics and Hillstream BioPharma. S. von Karstedt is named inventor on patent applications covering some of the therapeutic concepts pertaining to TRAIL-R blockade in disease. Y. H. Chen is a member of the board of advisors for Amshenn Inc. and Binde Inc. T.-D. Kanneganti is a consultant for Pfizer. T. Vanden Berghe and P. Vandenabeele hold patents related to ferrostatin-1 analogues. D. R. Green consults for Ventus Therapeutics, Inzen Therapeutics, and Horizon Therapeutics. K. H. Vousden is on the board of directors and shareholder of Bristol Myers Squibb and on the scientific advisory board (with stock options) of PMV Pharma, RAZE Therapeutics, Volastra Pharmaceuticals and Kovina Therapeutics. She is on the scientific advisory board of Ludwig Cancer and a co-founder and consultant of Faeth Therapeutics. She has been in receipt of research funding from Astex Pharmaceuticals and AstraZeneca and contributed to CRUK Cancer Research Technology filing of patent application WO/2017/144877. W. S. El-Deiry (WSE-D) is founder and shareholder (no research funding) of Oncoceutics/Chimerix, which is developing ONC201/TIC10 as cancer therapeutic. WSE-D is founder and shareholder (no research funding) of p53-Therapeutics, an early-stage company developing small molecules targeting mutant p53. WSE-D founder, and shareholder (no research funding) of SMURF-Therapeutics, an early-stage company developing small molecules targeting hypoxia. WSE-D is Co-Chair in the Executive Committee for Precision Oncology Alliance (no research funding) of Caris Life Sciences. WSE-D receives support from D&D Pharma and AACR-Novocure. WSE-D is the advisory board of Ocean Biomedical. WSE-D is the advisory board of RAIN Therapeutics. G. Kroemer has been holding research contra.cts with Daiichi Sankyo, Eleor, Kaleido, Lytix Pharma, PharmaMar, Osasuna Therapeutics, Samsara Therapeutics, Sanofi, Tollys, and Vascage. G. Kroemer has been consulting for Reithera. G. Kroemer is on the Board of Directors of the Bristol Myers Squibb Foundation France. G. Kroemer is a scientific co-founder of everImmune, Osasuna Therapeutics, Samsara Therapeutics and Therafast Bio. G. Kroemer is the inventor of patents covering therapeutic targeting of aging, cancer, cystic fibrosis and metabolic disorders. G. Kroemer’s brother, Romano Kroemer, was an employee of Sanofi and now consults for Boehringer-Ingelheim. L. Galluzzi is/has been holding research contracts with Lytix Biopharma, Promontory and Onxeo, has received consulting/advisory honoraria from Boehringer Ingelheim, AstraZeneca, OmniSEQ, Onxeo, The Longevity Labs, Inzen, Imvax, Sotio, Promontory, Noxopharm, EduCom, and the Luke Heller TECPR2 Foundation, and holds Promontory stock options. All other authors have no conflicts to declare.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Vitale, I., Pietrocola, F., Guilbaud, E. et al. Apoptotic cell death in disease—Current understanding of the NCCD 2023. Cell Death Differ 30, 1097–1154 (2023). https://doi.org/10.1038/s41418-023-01153-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41418-023-01153-w
This article is cited by
-
Targeting immunogenic cell stress and death for cancer therapy
Nature Reviews Drug Discovery (2024)
-
Targeting paraptosis in cancer: opportunities and challenges
Cancer Gene Therapy (2024)
-
Cancer cell metabolism and antitumour immunity
Nature Reviews Immunology (2024)
-
Diverse functions of cytochrome c in cell death and disease
Cell Death & Differentiation (2024)
-
Ferroptosis: Emerging mechanisms, biological function, and therapeutic potential in cancer and inflammation
Cell Death Discovery (2024)
