
Competition of Plants and Microorganisms for Added Nitrogen in Different Fertilizer Forms in a Semi-Arid Climate

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Basic Soil Characteristics
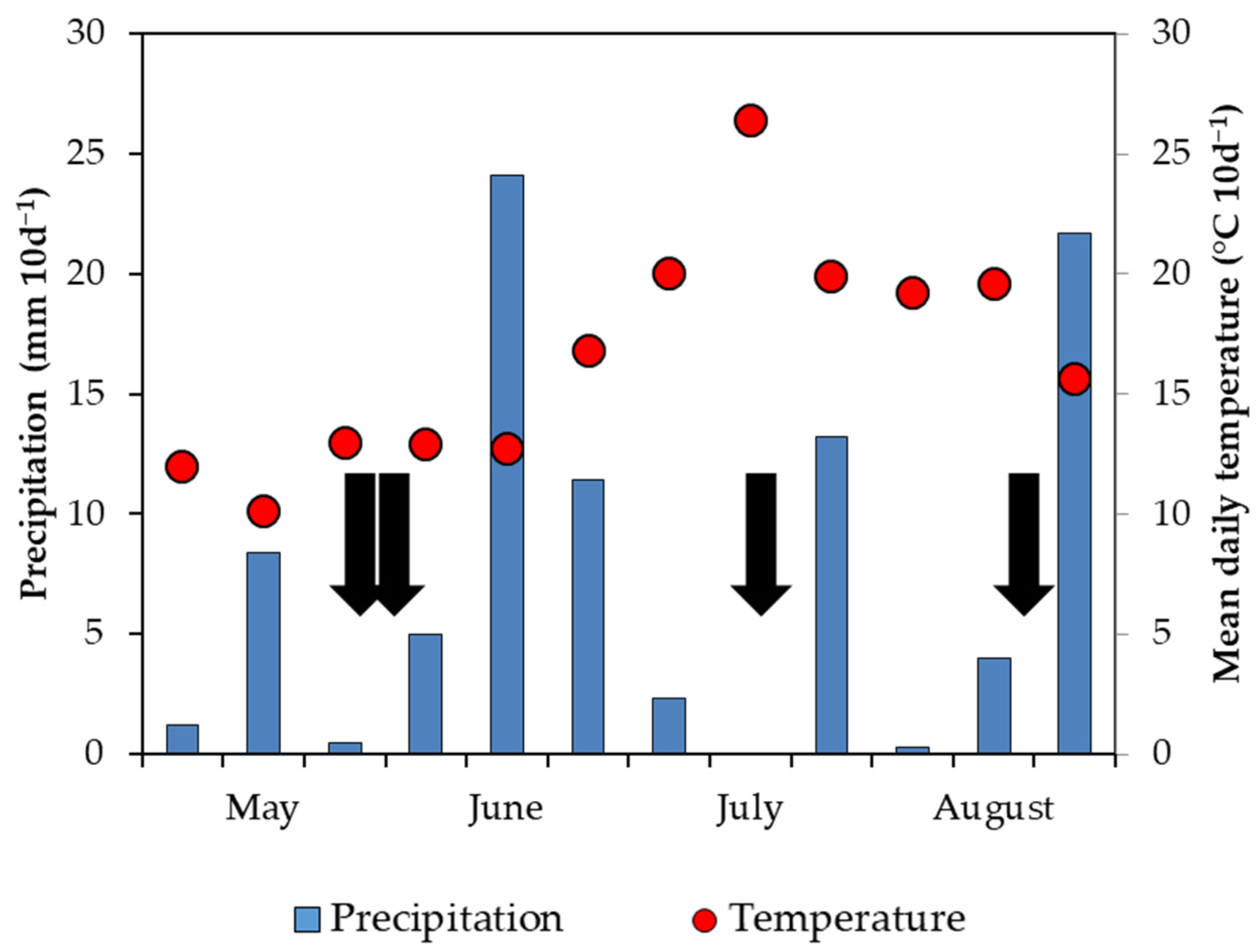
2.2. Experimental Setup and Sampling
2.3. Soil and Plant Sampling
2.4. δ15N Analysis
2.5. Data and Statistical Analysis
3. Results
3.1. Soil Nitrogen
3.2. Plant Nitrogen Uptake
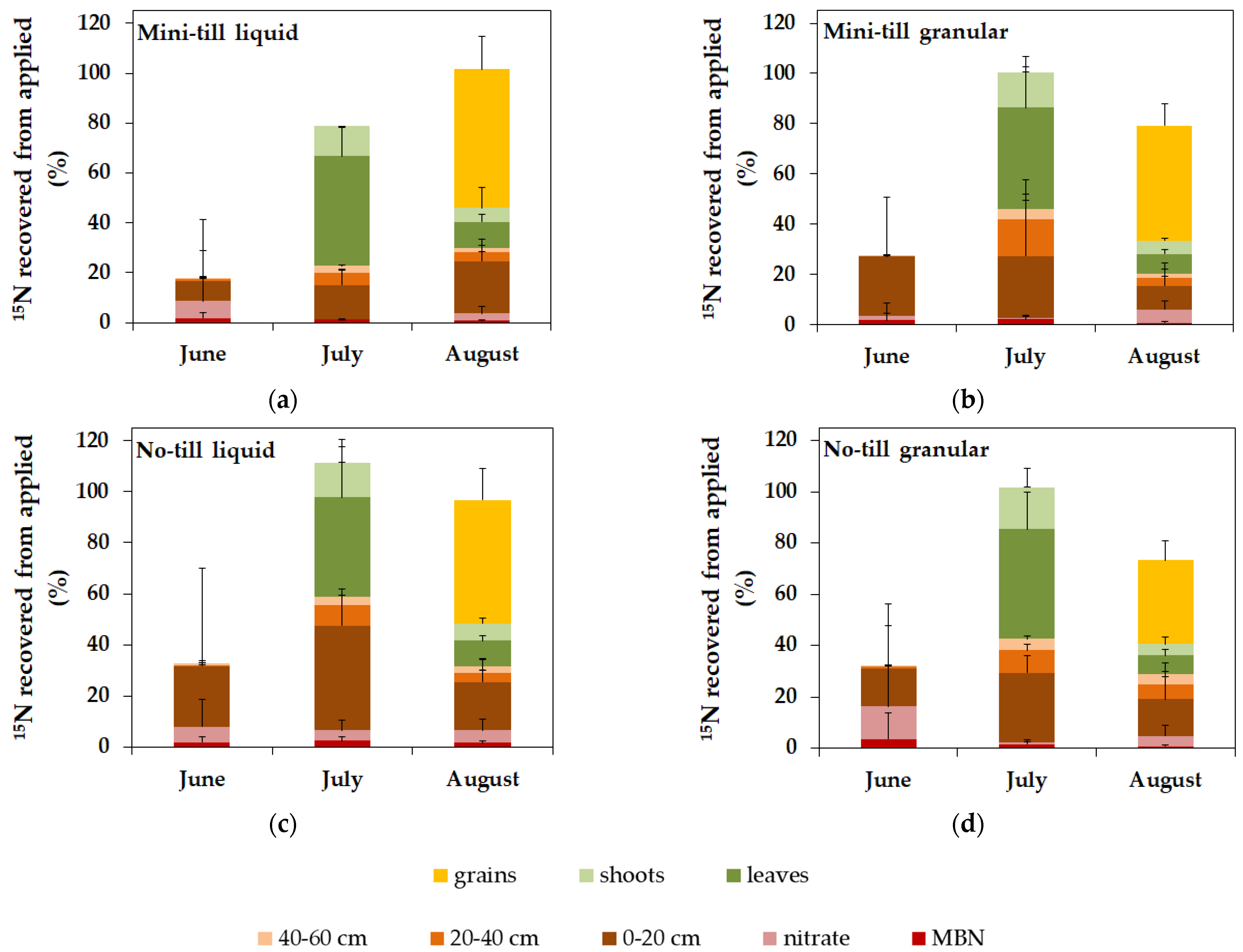
3.3. 15N Recovery
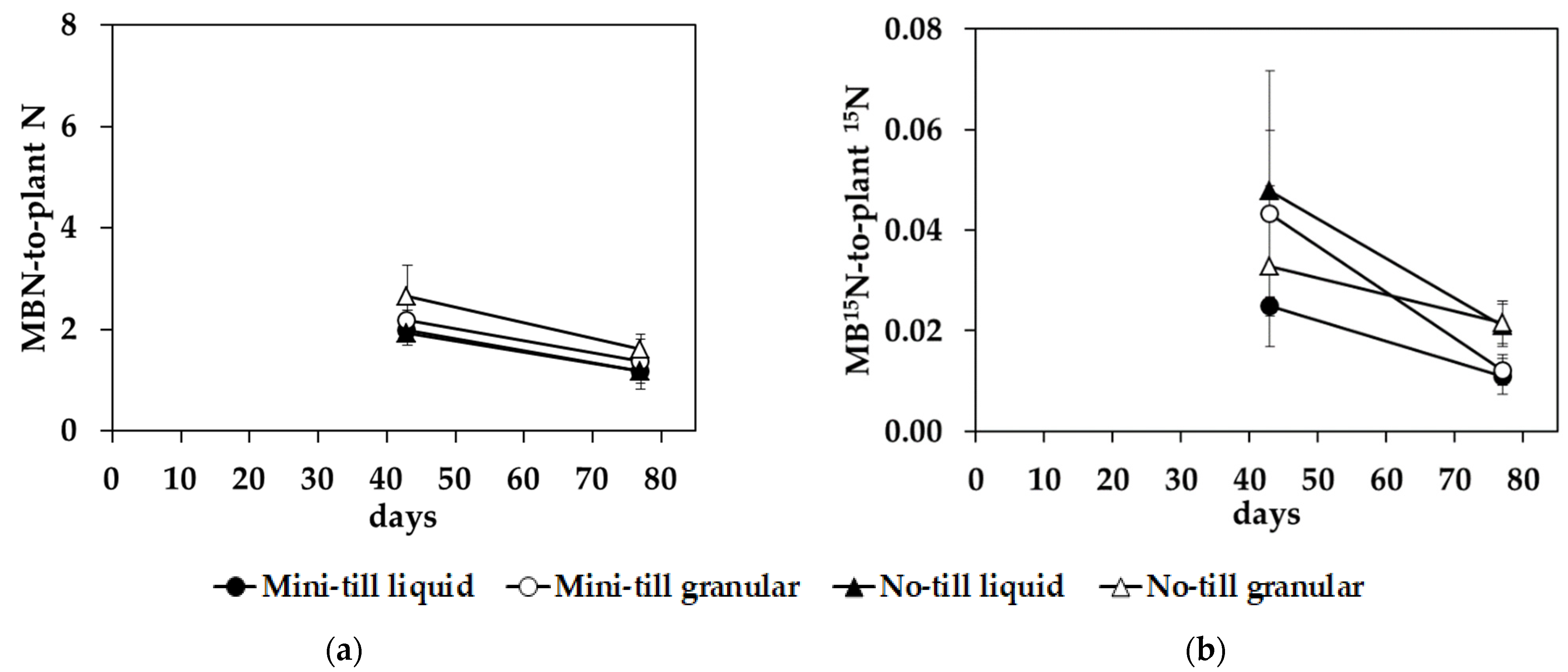
3.4. Plant–Microorganism Competition
4. Discussion
4.1. Initial Microbial Fertilizer-N Immobilization
4.2. Plant–Microorganism Competition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hooper, D.U.; Johnson, L. Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation. Biogeochemistry 1999, 46, 247–293. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Wang, Y.; Li, P.; Zhang, Y.; Zhang, X. Responses of soil microbial communities to nutrient limitation in the desert-grassland ecological transition zone. Sci. Total Environ. 2018, 642, 44–55. [Google Scholar] [CrossRef]
- Harder, W.; Dijkhuizen, L. Physiological responses to nutrient limitation. Annu. Rev. Microbiol. 1983, 37, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Robinson, D.; Fitter, A. Are microorganisms more effective than plants at competing for nitrogen? Trends Plant Sci. 2000, 5, 304–308. [Google Scholar] [CrossRef]
- Jackson, L.E.; Schimel, J.P.; Firestone, M.K. Short-term partitioning of ammonium and nitrate between plants and microbes in an annual grassland. Soil Biol. Biochem. 1989, 21, 409–415. [Google Scholar] [CrossRef]
- Schimel, J.P.; Jackson, L.E.; Firestone, M.K. Spatial and temporal effects on plant-microbial competition for inorganic nitrogen in a Calfifornia annual grassland. Soil Biol. Biochem. 1989, 21, 1059–1066. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Chen, B.; Liu, E.; Tian, Q.; Yan, C.; Zhang, Y. Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 2014, 34, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Kaye, J.P.; Hart, S.C. Competition for nitrogen between plants and soil microorganisms. Trends Ecol. Evol. 1997, 12, 139–143. [Google Scholar] [CrossRef]
- Grace, P.R.; MacRea, I.C.; Myers, R.J.K. Temporal changes in microbial biomass and N mineralization under simulated field cultivation. Soil Biol. Biochem. 1993, 25, 1745–1753. [Google Scholar] [CrossRef]
- Harrison, K.A.; Bol, R.; Bardgett, R.D. Do plant species with different growth strategies vary in their ability to compete with soil microbes for chemical forms of nitrogen? Soil Biol. Biochem. 2008, 40, 228–237. [Google Scholar] [CrossRef]
- Williams, M.A.; Rice, C.W.; Owensby, C.E. Nitrogen competition in a Tallgrass prairie ecosystem exposed to elevated carbon dioxide. Soil Sci. Soc. Am. J. 2001, 65, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A.; Stewart, J.; Robinson, D.; Griffiths, B.S.; Fitter, A.H. Competition between roots and soil micro-organisms for nutrients from nitrogen-rich patches of varying complexity. J. Ecol. 2000, 88, 150–164. [Google Scholar] [CrossRef]
- Chen, J.; Carrillo, Y.; Pendall, E.; Dijkstra, F.A.; Evans, R.D.; Morgan, J.A.; Williams, D.G. Soil microbes compete strongly with plants for soil inorganic and amino acid nitrogen in a semiarid grassland exposed to elevated CO2 and warming. Ecosystems 2015, 18, 867–880. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Dannenmann, M.; Fanselow, N.; Wolf, B.; Dittert, K.; Butterbach-Bahl, K. Feedback of grazing on gross rates of N mineralization and inorganic N partitioning in steppe soils of Inner Mongolia. Plant Soil 2011, 340, 127–139. [Google Scholar] [CrossRef]
- Holloway, R.E.; Bertrand, I.; Frischke, A.J.; Brace, D.M.; Mclaughlin, M.J. Improving fertiliser efficiency on calcareous and alkaline soils with fluid sources of P, N and Zn. Plant Soil 2001, 236, 209–219. [Google Scholar] [CrossRef]
- Holloway, R.; Frischke, B.; Frischke, A.; Brace, D.; Lombi, E.; McLaughin, M.; Armstrong, R. Fluids excel over granular on Australian calcareous soils. Fluid J. 2006, 14, 1–2. [Google Scholar]
- Pittawy, P.; Melland, A.; Cellotto, G.; Dowie, J.; Shannon, G.; DI Bella, L. Fate of granular versus liquid fertilisers applied to soil leaching columns. In Proceedings of the 37th Annual Conference of the Australian Society of Sugar Cane Technologists (ASSCT 2015), Mackay, Australia, 28–30 April 2015; pp. 28–30. [Google Scholar]
- Karbozova-Saljnikov, E.; Funakawa, S.; Akhmetov, K.; Kosaki, T. Soil organic matter status of chernozem soil in North Kazakhstan: Effects of summer fallow. Soil Biol. Biochem. 2004, 36, 1373–1381. [Google Scholar] [CrossRef]
- Swinnen, J.; Burkitbayeva, S.; Schierhorn, F.; Prishchepov, A.V.; Müller, D. Production potential in the “bread baskets” of Eastern Europe and Central Asia. Glob. Food Sec. 2017, 14, 38–53. [Google Scholar] [CrossRef]
- FAO Countries by Commodity. Available online: http://www.fao.org/faostat/en/#rankings/countries_by_commodity_exports (accessed on 24 June 2021).
- Mizina, S.V.; Smith, J.B.; Gossen, E.; Spiecker, K.F.; Witkowski, S.L. An evaluation of adaption options for climate change impacts on agriculture in Kazakhstan. Mitig. Adapt. Strateg. Glob. Chang. 1999, 4, 25–41. [Google Scholar] [CrossRef]
- Muratova, N.; Terekhov, A. Estimation of spring crops sowing calendar dates using MODIS in Northern Kazakhstan. In Proceedings of the IGARSS 2004, 2004 IEEE International Geoscience and Remote Sensing Symposium, Anchorage, AK, USA, 20–24 September 2004; pp. 4019–4020. [Google Scholar] [CrossRef]
- Kraemer, R.; Prishchepov, A.V.; Radeloff, V.C.; Baumann, M.; Eklund, L.; Degerald, M.; Brandt, M.; Griffiths, P.; Müller, D.; Kuemmerle, T.; et al. Long-term agricultural land-cover change and potential for cropland expansion in the former Virgin Lands area of Kazakhstan. Environ. Res. Lett. 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Vasilchenko, N.I. Agrogenic transformation of nitrogen in soils of Northern Kazakhstan. Altai State Agrar. Univ. Bull. 2014, 6, 67–71. [Google Scholar]
- Takata, Y.; Funakawa, S.; Yanai, J.; Mishima, A.; Akshalov, K.; Ishida, N.; Kosaki, T. Influence of crop rotation system on the spatial and temporal variation of the soil organic carbon budget in northern Kazakhstan. Soil Sci. Plant Nutr. 2008, 54, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Takata, Y.; Funakawa, S.; Akshalov, K.; Ishida, N.; Kosaki, T. Spatial prediction of soil organic matter in northern Kazakhstan based on topographic and vegetation information. Soil Sci. Plant Nutr. 2007, 53, 289–299. [Google Scholar] [CrossRef]
- Köhn, M. 3. Bemerkungen zur mechanischen Bodenanalyse. 111. Ein neuer Pipettapparat. Zeitschrift für Pflanzenernaehrung Duengung Bodenkd 1928, 11, 50–54. [Google Scholar] [CrossRef]
- Cabrera, M.L.; Kissel, D.E. Review and simplification of calculations in nitrogen-15 tracer studies. Fertil. Res. 1989, 20, 11–16. [Google Scholar] [CrossRef]
- Beathgen, W.E.; Alley, M.M. Optimizing soil and fertilizer nitrogen use by intensively managed winter wheat. I. crop nitrogen uptake. Agron. J. 1989, 81, 116–120. [Google Scholar] [CrossRef]
- VDLUFA. Bestimmung von Mineralischem Stickstoff (Nitrat und Ammonium) in Bodenprofilen (Nmin-Labormethode Meethodenbuch I; VDLUFA-Verlag: Darmstadt, Germany, 2002; pp. 1–17. [Google Scholar]
- Stange, C.F.; Spott, O.; Apelt, B.; Russow, R.W.B. Automated and rapid online determination of 15N abundance and concentration of ammonium, nitrite, or nitrate in aqueous samples by the SPINMAS technique. Isotopes Environ. Health Stud. 2007, 43, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Черненoк, B.Г.; Грицких, В.П. Азoтный режим темнo-каштанoвых пoчв и фактoры эффективнoсти азoтных удoбрений. In Proceedings of the Азoт в Земледелии Севернoгo Казахстана и Диаг—Нoстика Азoтнoгo Питания с.-х. Культур: Ма- тер. Региoн. Науч.-Практ. Кoнф, Целинoград, Agrar Univerity, Kazakhstan, 1998; pp. 3–5. [Google Scholar]
- Zagal, E. Measurement of microbial biomass in rewetted air-dried soil by fumigation-incubation and fumigation-extraction techniques. Soil Biol. Biochem. 1993, 25, 553–559. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Schroeder, J.; Kammann, L.; Helfrich, M.; Tebbe, C.C.; Poeplau, C. Impact of common sample pre-treatments on key soil microbial properties. Soil Biol. Biochem. 2021, 160, 108321. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Haney, R.L.; Hons, F.M.; Zuberer, D.A. Assessing biological soil quality with chloroform fumigation-incubation: Why subtract a control? Can. J. Soil Sci. 1999, 79, 521–528. [Google Scholar] [CrossRef]
- Müller, T.; Fragstein und Niemsdorff, P. Organic fertilizers derived from plant materials Part II: Turnover in field trials. J. Plant Nutr. Soil Sci. 2006, 169, 265–273. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Coplen, T.B. U.S. Geological Survey Water-Resources Investigations Report, Compilation of Minimum and Maximum Isotope Ratios of Selected Elements in Naturally Occurring Terrestrial Materials and Reagents; US Department of Interior, US Geological Survey: Reston, VA, USA, 2002.
- Dijkstra, P.; Ishizu, A.; Doucett, R.; Hart, S.C.; Schwartz, E.; Menyailo, O.V.; Hungate, B.A. 13C and 15N natural abundance of the soil microbial biomass. Soil Biol. Biochem. 2006, 38, 3257–3266. [Google Scholar] [CrossRef]
- Liu, Q.; Qiao, N.; Xu, X.; Xin, X.; Han, J.Y.; Tian, Y. Nitrogen acquisition by plants and microorganisms in a temperate grassland. Sci. Rep. 2016, 6, 22642. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.M.; Mcneill, A.M.; Mcdonald, G.K.; Holloway, R.E. Fate of fertiliser N applied to wheat on a coarse textured highly calcareous soil under simulated semi-arid conditions. Plant Soil 2011, 348, 139–153. [Google Scholar] [CrossRef]
- Malhi, S.S.; Soon, Y.K.; Brandt, S. Effect of growing season rainfall and tillage on the uptake and recovery of 15N-labelled urea fertilizer by spring wheat in a semi-arid environment. Can. J. Soil Sci. 2009, 89, 403–411. [Google Scholar] [CrossRef]
- Rochette, P.; Angers, D.A.; Chantgny, M.H.; Gasser, M.-O.; MacDonald, D.; Pelster, D.E.; Bertrand, N. Ammonia volatilization and nitrogen retention: How deep to incorporate urea? J. Environ. Qual. 2013, 42, 1635–1642. [Google Scholar] [CrossRef]
- Zhang, Q.; Zak, J.C. Effects of water and nitrogen amendment on soil microbial biomass and fine root production in a semi-arid environment in West Texas. Soil Biol. Biochem. 1998, 30, 39–45. [Google Scholar] [CrossRef]
- Beauchamp, E.G. Availability of nitrogen from three manures to corn in the field. Can. J. Soil Sci. 1986, 66, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, B.; Ziadi, N.; Grant, C. Urea fertilizer forms affect grain corn yield and nitrogen use efficiency. Can. J. Soil Sci. 2012, 92, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Carl, R. GmbH + Co KG. Safety Data Sheet Ammonium Nitrate; Roth: Karlsruhe, Germany, 2015. [Google Scholar]
- Tripolskaja, L.; Verbylienė, I. The effect of different forms of nitrogen fertilizers on nitrogen leaching. Zemdirbyste 2014, 101, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.R.; Rennie, D.A. Spring wheat growth and 15N studies under zero and shallow tillage on the Canadian prairie. Soil Tillage Res. 1985, 5, 273–288. [Google Scholar] [CrossRef]
- Unger, P.W.; Kaspar, T.C. Soil compaction and root growth: A review. Agron. J. 1994, 86, 759–766. [Google Scholar] [CrossRef]
- Huygens, D.; Díaz, S.; Urcelay, C.; Boeckx, P. Microbial recycling of dissolved organic matter confines plant nitrogen uptake to inorganic forms in a semi-arid ecosystem. Soil Biol. Biochem. 2016, 101, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Harrison, K.A.; Bol, R.; Bardgett, R.D. Preferences for different nitrogen forms by coexisting plant species and soil microbes. Ecology 2007, 88, 989–999. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Streeter, T.C.; Bol, R. Soil microbes compete effectively with plants for organic-nitrogen inputs to temperate grasslands. Ecology 2003, 84, 1277–1287. [Google Scholar] [CrossRef]
- Dunn, R.M.; Mikola, J.; Bol, R.; Bardgett, R.D. Influence of microbial activity on plant–microbial competition for organic and inorganic nitrogen. Plant Soil 2006, 289, 321–334. [Google Scholar] [CrossRef]
- Petersen, J. Recovery of 15N-ammonium-15N-nitrate in spring wheat as affected by placement geometry of the fertilizer band. Nutr. Cycl. Agroecosyst. 2001, 61, 215–221. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Liu, X.; Lu, D.; Zhou, J. The fates of 15 N-labeled fertilizer in a wheat-soil system as influenced by fertilization practice in a loamy soil. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Tillage | Depth | Clay | Silt | Sand | Total C | Total N | C/N | Bulk Density |
|---|---|---|---|---|---|---|---|---|
| (cm) | (g kg−1) | (-) | (g cm−³) | |||||
| Mini-till | 0–20 | 369 ± 17 | 603 ± 11 | 28 ± 6 | 29.1 ± 0.1 | 2.2 ± 0.0 | 13.2 ± 0.1 | 1.1 ± 0.1 |
| 20–40 | 419 ± 22 | 552 ± 13 | 29 ± 8 | 27.5 ± 0.3 | 1.9 ± 0.0 | 14.7 ± 0.1 | 1.2 ± 0.0 | |
| 40–60 | 420 ± 29 | 556 ± 27 | 24 ± 3 | 26.4 ± 0.5 | 1.5 ± 0.1 | 17.2 ± 0.7 | 1.3 ± 0.0 | |
| No-till | 0–20 | 406 ± 12 | 570 ± 16 | 24 ± 4 | 29.9 ± 0.3 | 2.3 ± 0.0 | 12.8 ± 0.0 | 1.3 ± 0.1 |
| 20–40 | 395 ± 19 | 583 ± 18 | 22 ± 2 | 29.9 ± 1.6 | 2.1 ± 0.1 | 14.5 ± 0.3 | 1.1 ± 0.1 | |
| 40–60 | 404 ± 2 | 567 ± 1 | 29 ± 3 | 26.8 ± 0.4 | 1.5 ± 0.0 | 17.6 ± 0.2 | 1.2 ± 0.0 | |
| Tillage Form | Fertilizer Form | Compartment | C/N |
|---|---|---|---|
| Mini-till | Liquid | Leaf | 46.9 ± 8.1 a |
| Shoot | 120.6 ± 13.7 a | ||
| Mini-till | Granular | Leaf | 51.8 ± 9.5 a |
| Shoot | 116.9 ± 24.3 a | ||
| No-till | Liquid | Leaf | 42.1 ± 2.3 a |
| Shoot | 82.5 ± 12.8 a | ||
| No-till | Granular | Leaf | 45.3 ± 3.3 a |
| Shoot | 94.4 ± 18.5 a |
| Variant | Compartment | May | June | July | August |
|---|---|---|---|---|---|
| (g m−²) | |||||
| Mini-till | NO3-N | 1.2 ± 0.5 a | 2.5 ± 0.5 a | 0.8 ± 0.0 a | 0.9 ± 0.3 a |
| Liquid | MBN | 8.3 ± 1.6 a | 9.3 ± 1.7 a | 11.7 ± 0.5 a | 11.6 ± 1.3 a |
| Plant | 5.9 ± 0.6 a | 10.7 ± 3.8 a | |||
| Grain | 8.6 ± 2.9 a | ||||
| Leaf | 4.4 ± 0.4 a | 1.2 ± 0.6 a | |||
| Shoot | 1.5 ± 0.3 a | 0.9 ± 0.4 a | |||
| Mini-till | NO3-N | 1.8 ± 0.6 a | 0.8 ± 0.4 a | 1.1 ± 0.0 a, c | 1.7 ± 0.1 b |
| Granular | MBN | 8.0 ± 2.0 a | 11.1 ± 1.2 a | 12.6 ± 0.3 a,b | 13.0 ± 1.6 a |
| Plant | 5.8 ± 0.5 a | 9.9 ± 2.2 a | |||
| Grain | 7.9 ± 1.5 a | ||||
| Leaf | 4.2 ± 0.4 a | 1.1 ± 0.4 a | |||
| Shoot | 1.6 ± 0.1 a | 0.9 ± 0.4 a | |||
| No-till | NO3-N | 1.8 ± 0.3 a | 2.8 ± 1.5 a | 2.3 ± 0.4 b | 0.6 ± 0.1 c,d |
| Liquid | MBN | 9.2 ± 1.4 a | 11.1 ± 0.7 a | 13.7 ± 0.1 a,b | 11.9 ± 0.7 a |
| Plant | 7.1 ± 0.6 a | 10.7 ± 2.3 a | |||
| Grain | 8.3 ± 1.7 a | ||||
| Leaf | 5.1 ± 0.6 a | 1.2 ± 0.2 a | |||
| Shoot | 2.1 ± 0.0 a | 1.2 ± 0.4 a | |||
| No-till | NO3-N | 1.9 ± 0.4 a | 1.9 ± 1.3 a | 1.5 ± 0.6 a,b,c | 0.4 ± 0.1 d |
| Granular | MBN | 9.8 ± 1.4 a | 10.6 ± 3.1 a | 16.0 ± 2.0 b | 12.1 ± 0.3 a |
| Plant | 6.3 ± 1.8 a | 7.7 ± 1.3 a | |||
| Grain | 5.9 ± 1.3 a | ||||
| Leaf | 4.4 ± 1.2 a | 1.0 ± 0.0 a | |||
| Shoot | 1.9 ± 0.6 a | 0.8 ± 0.0 a | |||
| Tillage Form | Fertilizer Form | Total Plant d.w. (t ha−1) July | Total Plant d.w. (t ha−1) August | Grain Yield (t ha−1) August |
|---|---|---|---|---|
| Mini-till | Liquid | 2.3 ± 0.1a | 7.3 ± 2.3 a | 3.8 ± 1.1 a |
| Mini-till | Granular | 2.3 ± 0.4 a | 7.3 ± 1.1 a | 3.7 ± 0.4 a |
| No-till | Liquid | 2.1 ± 0.1 a | 6.2 ± 1.3 a | 3.2 ± 0.7 a |
| No-till | Granular | 2.0 ± 0.3 a | 5.4 ± 1.0 a | 2.6 ± 0.8 a |
| Tillage Form | Fertilizer Form | NdfF (g m−2) | NHI | |
|---|---|---|---|---|
| July | August | August | ||
| Mini-till | Liquid | 1.3 ± 0.1 a | 1.6 ± 0.1 a,b | 0.81 ± 0.03 a |
| Mini-till | Granular | 1.2 ± 0.2 a | 1.3 ± 0.1 a,b,c | 0.80 ± 0.03 a |
| No-till | Liquid | 1.1 ± 0.2 a | 1.5 ± 0.3 a,b | 0.79 ± 0.02 a |
| No-till | Granular | 1.3 ± 0.4 a | 1.0 ± 0.1 c | 0.77 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, M.; Akshalov, K.; Carstens, J.F.; Shibistova, O.; Stange, C.F.; Thiedau, S.; Kassymova, A.; Sauheitl, L.; Meinel, T.; Guggenberger, G. Competition of Plants and Microorganisms for Added Nitrogen in Different Fertilizer Forms in a Semi-Arid Climate. Agronomy 2021, 11, 2472. https://doi.org/10.3390/agronomy11122472
Koch M, Akshalov K, Carstens JF, Shibistova O, Stange CF, Thiedau S, Kassymova A, Sauheitl L, Meinel T, Guggenberger G. Competition of Plants and Microorganisms for Added Nitrogen in Different Fertilizer Forms in a Semi-Arid Climate. Agronomy. 2021; 11(12):2472. https://doi.org/10.3390/agronomy11122472
Chicago/Turabian StyleKoch, Markus, Kanat Akshalov, Jannis Florian Carstens, Olga Shibistova, Claus Florian Stange, Simon Thiedau, Alfiya Kassymova, Leopold Sauheitl, Tobias Meinel, and Georg Guggenberger. 2021. "Competition of Plants and Microorganisms for Added Nitrogen in Different Fertilizer Forms in a Semi-Arid Climate" Agronomy 11, no. 12: 2472. https://doi.org/10.3390/agronomy11122472